1. 蛙類簡介
蛙類是一群極具特色的兩棲動物,在地球上已存在超過2億年。牠們的身體結構適應了水陸兩棲的生活方式,具有光滑濕潤的皮膚、發達的後肢和無尾的身軀。蛙類在分類學上屬於無尾目(Anura),是兩棲綱中最大的一個目,全球已知約有7000多種。
蛙類在生態系統中扮演著重要的角色。作為食物鏈中的中間環節,牠們既是許多捕食者的獵物,也是控制昆蟲和其他小型動物數量的重要力量。此外,蛙類對環境變化極為敏感,常被科學家用作生態系統健康狀況的指標物種。
在人類文化中,蛙類也占有獨特的地位。在許多文化傳統中,蛙被視為多產、好運或變革的象徵。例如,在中國文化中,「蛙」的諧音與「發」相近,因此常被視為財富和好運的象徵。在西方文化中,青蛙王子的童話故事廣為流傳,反映了人們對蛙類的特殊情感。
蛙類的生活史十分獨特,大多數種類經歷由水生蝌蚪到陸生成蛙的變態過程,這種巨大的形態和生理變化一直是生物學研究的熱點。同時,蛙類多樣的繁殖策略也引人注目,從水中產卵到背負幼蛙,展現了驚人的適應能力。
儘管蛙類在生態和文化中如此重要,但當前面臨著諸多威脅,如棲息地破壞、氣候變遷和疾病等。因此,加強對蛙類的研究和保護不僅對維護生物多樣性至關重要,也關乎人類自身的福祉。
1.1 什麼是蛙類?
蛙類是一群獨特而多樣的兩棲動物,屬於脊椎動物門中的兩棲綱無尾目。這個龐大的類群包括了青蛙、蟾蜍以及樹蛙等,全球已知約有7,000多種。蛙類的演化歷史可以追溯到2.65億年前的二疊紀晚期,經過漫長的時間演化,適應了各種不同的生態環境。
蛙類的身體結構具有許多特徵性的適應。它們通常擁有一個扁平的身體,沒有明顯的頸部,頭部與軀幹相連。大多數蛙類都沒有尾巴,這也是「無尾目」名稱的由來。蛙類的後肢特別發達,適合跳躍和游泳。這些強壯的後肢使得某些種類能夠跳躍超過自身體長20倍的距離。例如,古巴樹蛙(Osteopilus septentrionalis)能夳跳躍超過1.5米的距離。
蛙類的皮膚是其最顯著的特徵之一。它們擁有光滑、濕潤且高度通透的皮膚,不僅用於呼吸,還能夠吸收水分和某些養分。這種獨特的皮膚結構使得蛙類能夠在陸地上生存,同時保持體內水分平衡。然而,這也使得蛙類對環境變化和污染極為敏感。例如,酸雨會直接通過皮膚影響蛙類的生理功能,導致種群數量下降。
在感官系統方面,蛙類擁有突出的大眼睛,視力優秀,能夠捕捉快速移動的獵物。它們的眼睛還具有特殊的瞬膜,可以在水下保護眼睛。蛙類的聽覺同樣發達,耳朵位於眼睛後方,能夠接收空氣和水中的聲波。許多種類還具有鼓膜,有助於增強聽覺能力。例如,中國雨蛙(Hyla chinensis)能夠精確地定位50米外的聲源。
蛙類的生活史是其另一個引人注目的特點。大多數蛙類經歷複雜的變態過程,從水生的蝌蚪階段轉變為陸生的成體。這個過程涉及到全身器官系統的重組,是生物學研究中的重要課題。一些特殊的蛙類,如智利袋蟾(Rhinoderma darwinii),甚至進化出了獨特的繁殖方式,雄性會將受精卵存放在自己的發聲囊中孵化。
在生態系統中,蛙類扮演著關鍵的角色。作為食物鏈中的中間環節,它們既是許多捕食者的獵物,也是控制昆蟲和其他小型無脊椎動物數量的重要力量。例如,一隻成年的美洲牛蛙(Lithobates catesbeianus)每年可以消耗數千隻昆蟲,對農業害蟲的控制起到重要作用。同時,蛙類的蝌蚪在水域生態系統中也扮演著重要角色,它們通過攝食藻類和有機碎屑來調節水質。
蛙類的多樣性體現在它們的棲息地選擇上。從熱帶雨林到沙漠,從高山到平原,幾乎所有的陸地生態系統都能找到蛙類的蹤跡。有些種類,如澳大利亞綠樹蛙(Litoria caerulea),幾乎整個生命週期都在樹上度過;而非洲爪蟾(Xenopus laevis)則完全適應了水生環境,終生生活在水中。這種多樣的棲息地選擇反映了蛙類驚人的適應能力。
蛙類在人類文化中也占有重要地位。在許多文化傳統中,蛙被視為多產、好運或變革的象徵。例如,在中國文化中,「蛙」的諧音與「發」相近,因此常被視為財富和好運的象徵。青蛙玉蟾常被用作裝飾品,象徵著財源廣進。在古埃及,蛙神黑卡特(Heqet)是生育與重生的象徵,與尼羅河的泛濫和新生命的誕生相聯繫。
在現代科學研究中,蛙類也發揮著重要作用。它們常被用作模式生物,用於研究發育生物學、生態學和毒理學等領域。例如,非洲爪蟾因其易於飼養和基因組簡單,成為了發育生物學研究中的重要模式生物。此外,蛙類皮膚分泌物中的生物活性物質也為新藥研發提供了豐富的資源。南美箭毒蛙(Dendrobates)皮膚分泌物中的生物鹼已被用於開發新型鎮痛藥。
然而,儘管蛙類如此重要,它們目前正面臨著前所未有的威脅。棲息地破壞、氣候變化、環境污染、外來物種入侵以及新興疾病如壺菌病(Chytridiomycosis)等,都對全球蛙類種群造成了嚴重影響。據國際自然保護聯盟(IUCN)的數據,目前約有40%的兩棲類物種面臨滅絕威脅,其中很大一部分是蛙類。
面對這些挑戰,科學家們正在採取各種措施來保護蛙類。這包括棲息地保護、人工繁殖瀕危物種、疾病防控等多方面的努力。例如,為了挽救瀕臨滅絕的巴拿馬金蛙(Atelopus zeteki),科學家們建立了人工飼養計劃,希望能在將來重新將其引入野外。同時,公眾教育和參與也在蛙類保護中扮演著重要角色,越來越多的人開始關注並參與到蛙類及其棲息地的保護工作中。
1.2 蛙類分類學概述
蛙類的分類學是生物學中一個充滿挑戰和不斷演變的領域。為兩棲綱中最大的一個目,分佈在全球各個大陸,除了南極洲。蛙類的分類體系經過了長期的發展和多次修訂,反映了我們對這些生物認識的不斷深入。
傳統上,蛙類的分類主要基於形態學特徵。早期的分類學家主要依靠外部形態、骨骼結構和內部解剖特徵來區分不同的種類。例如,蛙類和蟾蜍曾經被認為是兩個截然不同的群體,主要基於它們的皮膚質地和腿部肌肉的差異。然而,隨著研究的深入,我們發現這種簡單的二分法並不能準確反映蛙類的演化關係。
現代蛙類分類學結合了形態學、分子生物學、生態學和行為學等多個學科的研究方法。分子生物學技術的發展,特別是DNA序列分析的應用,極大地改變了我們對蛙類系統發育關係的理解。例如,通過基因序列分析,科學家發現了一些外表相似但實際上並不密切相關的蛙類,這種現象被稱為趨同演化。反之,一些形態差異很大的種類可能實際上是近親。
目前,無尾目被劃分為多個科(Fami)。其中一些主要的科包括:蛙科(Ranidae)、樹蛙科(Hylidae)、蟾蜍科(Bufonidae)、狹口蛙科(Microhylidae)等。每個科又進一步分為屬(Genus)和種(物種)。值得注意的是,隨著新物種的發現和分子生物學研究的深入,這些分類群的界定和關係常常會發生變化。
蛙科是最大的一個科,包含了眾多常見的蛙類。這個科的成員通常具有長而有力的後腿,適合跳躍和游泳。青蛙屬(Rana)是蛙科中最知名的屬之一,包括了許多廣泛分佈的物種。例如,在台灣常見的長腳赤蛙(Rana longicrus)就屬於這個屬。
樹蛙科的成員通常具有特化的吸盤腳趾,使它們能夠在樹上和其他垂直表面活動。這個科的物種通常具有鮮豔的色彩和優雅的體型。台灣特有種諸羅樹蛙(Rhacophorus prasinatus)就是一個典型的例子,它能夠在樹冠層中靈活移動。
蟾蜍科的成員通常體型較為粗壯,皮膚上有明顯的疣狀突起。這些突起實際上是毒腺,能夠分泌有毒物質以抵禦捕食者。海蟾蜍(Rhinella marina)是這個科中最知名的物種之一,由於其強大的適應能力,在被引入澳大利亞等地後成為了危險的入侵物種。
狹口蛙科的成員通常體型較小,頭部呈三角形,口部狹小。這個科的許多物種具有特殊的生活習性,如穴居或半穴居。小雨蛙(Microhyla fissipes)是台灣常見的狹口蛙科物種,其獨特的鳴叫聲是夏夜的特色之一。
除了這些大型的科,還有許多較小但同樣重要的科。例如,箭毒蛙科(Dendrobatidae)的成員以其鮮豔的警戒色彩和強烈的毒性而聞名。角蛙科(Ceratophryidae)的成員則以其巨大的口部和獨特的形態引人注目。
蛙類的分類學還面臨著許多挑戰。隱蔽種(Cryptic 物種)的存在是其中之一。這些物種在形態上幾乎無法區分,但實際上是獨立的演化支系。例如,在歐洲的綠蛙複合體(Pelophylax esculentus complex)中,就包含了多個形態相似但遺傳上截然不同的物種。這種情況需要結合形態學、聲學和分子生物學等多種方法才能準確分類。
另一個挑戰是物種界定的問題。隨著我們對基因流動和雜交現象認識的深入,傳統的生物學物種概念(Biological 物種 Concept)在某些情況下顯得不夠完善。例如,某些蛙類群體之間存在有限的基因交流,但仍保持了明顯的形態和生態差異。這種情況下,如何界定物種邊界就成為了一個複雜的問題。
蛙類的分類學還與保護生物學密切相關。準確的分類信息對於制定有效的保護策略至關重要。例如,台灣的梭德氏赤蛙(Rana sauteri)原本被認為是廣泛分佈的物種,但近年的研究表明它實際上是台灣的特有種。這樣的發現直接影響了相關的保護措施。
此外,新物種的發現也不斷豐富著蛙類的分類系統。即使在21世紀,科學家們仍在世界各地發現新的蛙類物種。例如,2019年在印度西高止山脈發現的紫色蛙(Nasikabatrachus bhupathi)不僅是一個新物種,還代表了一個新的屬。這些發現不僅增加了我們對生物多樣性的認識,也為蛙類的演化史提供了新的線索。
隨著研究方法的進步,特別是基因組學和生物信息學技術的發展,蛙類的分類學正在經歷一場革命。這些新技術使我們能夠更準確地重建蛙類的演化樹,了解不同群體之間的親緣關係。然而,這也帶來了新的挑戰,如如何整合形態學和分子數據,以及如何處理大量的基因組數據。
蛙類的分類學是一個動態的領域,反映了我們對這些生物認識的不斷深入。它不僅是一門純粹的學術研究,還與生態保護、進化生物學等多個領域密切相關。隨著研究的不斷深入,我們對蛙類多樣性的認識也在不斷豐富和完善。
1.3 蛙類演化的重要性
蛙類在演化生物學中占有極其重要的地位,它們的演化歷程不僅反映了地球生命史上的重大轉折,還為我們理解脊椎動物的適應性輻射提供了寶貴的案例。蛙類的祖先大約在泥盆紀晚期到石炭紀早期(約3.7億年前)從魚類演化而來,成為最早適應陸地生活的脊椎動物之一。這次從水生到陸生的轉變標誌著脊椎動物演化史上的一個重要里程碑。
蛙類的演化過程中最引人注目的特徵之一是它們獨特的變態過程。從水生的蝌蚪到陸生的成體,這種戲劇性的形態變化不僅體現了個體發育過程中的重大轉變,也反映了蛙類祖先從水生到陸生的演化歷程。這種變態過程涉及全身器官系統的重組,包括呼吸系統從鰓到肺的轉變、循環系統的重構、以及骨骼和肌肉系統的重塑。研究蛙類的變態過程為我們理解脊椎動物早期演化提供了重要線索。
蛙類的骨骼結構演化也極具意義。與其他四足動物相比,蛙類的骨骼結構經歷了顯著的簡化和特化。例如,蛙類的脊椎骨數量大幅減少,尾椎融合成一個單一的尾骨(尾杆骨)。這種結構使蛙類能夠進行有效的跳躍運動。同時,蛙類的頭骨也經歷了顯著的改變,形成了獨特的扁平結構,有利於捕捉獵物和吞嚥。這些骨骼變化展示了生物如何通過結構簡化來實現功能的優化,為我們理解形態演化提供了重要案例。
蛙類的皮膚結構也是其演化重要性的一個體現。與其他陸生脊椎動物不同,蛙類保留了高度通透的皮膚,能夠通過皮膚進行呼吸和水分交換。這種特性使蛙類能夠在不同的環境中生存,但同時也使它們對環境變化特別敏感。蛙類皮膚的演化為我們理解生物與環境互動的機制提供了獨特的視角。
在繁殖策略方面,蛙類展現了令人驚嘆的多樣性,反映了它們適應不同環境的能力。從最常見的水中產卵到一些特殊的繁殖方式,如背部孵卵、口腔孵卵等,蛙類的繁殖策略涵蓋了脊椎動物繁殖模式的廣泛範圍。例如,智利袋蟾(Rhinoderma darwinii)的雄性會將受精卵吞入口腔,在特化的聲囊中孵化幼蛙。這種獨特的繁殖方式展示了生物為適應特定環境所進行的極端特化。
蛙類在聲音通訊方面的演化也極具意義。作為最早發展出有效聲音通訊系統的陸生脊椎動物之一,蛙類的鳴叫行為為我們理解動物通訊的演化提供了重要線索。不同種類的蛙類發展出了各式各樣的發聲器官和鳴叫模式,這些差異不僅反映了物種間的隔離機制,也展示了聲音信號在動物行為中的重要作用。
在生態適應方面,蛙類展現了驚人的多樣性。從熱帶雨林到乾旱沙漠,從高山到平原,蛙類幾乎適應了地球上所有的陸地生態系統。這種廣泛的分佈反映了蛙類強大的適應能力和演化潛力。例如,某些沙漠蛙類能夠長期埋在沙中度過乾旱期,而一些高山蛙類則進化出了抗凍機制來應對嚴寒環境。這些適應性特徵不僅展示了生物演化的多樣性,也為我們理解氣候變化對生物的影響提供了重要參考。
蛙類在毒素演化方面也提供了獨特的研究案例。許多蛙類,特別是新熱帶區的箭毒蛙,進化出了複雜的毒素系統。這些毒素不僅作為防禦機制,還在某些情況下被土著人用作狩獵工具。研究這些毒素的演化過程和作用機制,為我們了解生物化學防禦系統的發展提供了寶貴的線索,同時也為新藥研發開闢了新的途徑。
在分子演化研究中,蛙類也扮演著重要角色。由於蛙類種類繁多,分佈廣泛,它們成為研究分子鐘和生物地理學的理想對象。通過比較不同蛙類物種的基因序列,科學家們能夠重建它們的演化歷史,了解物種形成和遷移的過程。例如,通過研究非洲和南美洲蛙類的親緣關係,科學家們獲得了支持大陸漂移理論的重要證據。
蛙類的演化還為我們理解快速適應性輻射提供了絕佳的案例。在某些isolated的環境中,如馬達加斯加島,蛙類展現了驚人的多樣化。短時間內形成大量不同形態和生態位的物種,這種現象為我們理解物種形成的機制提供了重要線索。
此外,蛙類在研究氣候變化和環境污染對生物的影響方面也發揮著關鍵作用。由於其特殊的生理特征和生活史特点,蛙類對環境變化特別敏感。全球範圍內蛙類種群的下降被視為生態系統健康狀況的重要指標。研究蛙類對環境變化的響應,有助於我們預測和理解全球變化對生態系統的影響。
蛙類的演化重要性還體現在它們與其他生物的協同演化關係上。例如,某些蛙類與特定植物之間形成了互利共生關係,蛙類在植物的葉片間隙中築巢,同時為植物提供養分。這種關係的演化過程展示了生物間相互作用如何塑造生態系統。
1.4 蛙類的生態角色
蛙類在生態系統中扮演著多重而關鍵的角色,它們的存在和活動對於維持生態平衡至關重要。作為兩棲動物,蛙類在水陸生態系統之間架起了一座重要的橋樑,將這兩個環境緊密聯繫在一起。
首先,蛙類是食物網中的重要一環。作為中間消費者,蛙類既是捕食者,又是被捕食者。在幼體階段,蝌蚪主要以水生植物、藻類和有機碎屑為食,在水域生態系統中扮演著初級消費者的角色。這一過程有助於控制水體中的藻類數量,維持水質平衡。例如,在某些淡水生態系統中,蝌蚪的存在可以顯著減少水體優養化的風險。
當蛙類發育成成體後,它們成為了重要的昆蟲捕食者。一隻成年蛙類每天可以消耗大量的昆蟲,包括蚊子、蒼蠅和其他可能傳播疾病的昆蟲。在農業生態系統中,蛙類對於控制害蟲數量具有重要作用。例如,在亞洲的水稻田中,水蛙(Rana limnocharis)被認為是控制稻田害蟲的天然盟友。這種生物防治作用不僅有助於減少農藥使用,還能維持生態系統的自然平衡。
同時,蛙類也是許多掠食者的重要食物來源。鳥類、蛇類、魚類,甚至一些哺乳動物都以蛙類為食。這使得蛙類成為將水生生態系統的能量轉移到陸地生態系統的重要媒介。例如,在北美的某些濕地生態系統中,遷徙鳥類在繁殖季節大量捕食蛙類,這種能量轉移對於維持更廣泛的生態網絡至關重要。
蛙類在養分循環中也發揮著重要作用。當蛙類在水域中產卵和孵化時,未能發育的卵和死亡的蝌蚪成為水生生物的養分來源。同樣,當成年蛙類死亡時,它們的屍體為陸地生態系統提供了養分。這種養分循環對於維持生態系統的健康和穩定性具有重要意義。
蛙類還是生態系統健康狀況的重要指標。由於其特殊的生理特徵,如半透膜的皮膚,蛙類對環境變化極為敏感。水質污染、氣候變化、紫外線輻射增強等環境因素都會對蛙類產生顯著影響。因此,蛙類種群的變化常常被用作評估生態系統健康狀況的重要參考。例如,在北美的某些地區,科學家們通過監測斑點樹蛙(Hyla versicolor)的種群數量來評估森林生態系統的整體健康狀況。
在某些生態系統中,蛙類還扮演著生態工程師的角色。例如,在亞馬遜雨林中,樹蛙的存在能夠改變樹冠層的微環境。這些樹蛙在樹葉上產卵,形成小型的水池。這些微小的水體不僅為其他生物提供了棲息地,還改變了局部的濕度和養分分佈,進而影響整個生態系統的功能。
蛙類的繁殖活動也對生態系統產生重要影響。大量蛙類的同步產卵行為可以在短時間內為水域生態系統提供大量的有機物質,這不僅為水生生物提供了食物來源,還影響了水體的化學特性。例如,在某些季節性池塘中,蛙類的集體產卵活動可以顯著提高水體的氮磷含量,進而影響整個水域生態系統的動態平衡。
蛙類還在種子傳播方面發揮著作用。雖然不如鳥類和哺乳動物那樣顯著,但一些研究表明,某些蛙類可以通過消化系統傳播小型種子。這種傳播方式雖然範圍有限,但在局部生態系統中可能產生重要影響,尤其是對於一些濕地植物的傳播。
在一些特殊的生態系統中,蛙類還與其他生物形成了獨特的共生關係。例如,在中美洲的某些地區,金色箭毒蛙(Dendrobates auratus)與特定的樹種形成了互利共生關係。這種蛙類在樹洞中產卵和撫育幼蛙,同時它們的排泄物為樹木提供了養分。這種關係不僅體現了生態系統中物種間複雜的相互依存,還展示了蛙類在維持生物多樣性方面的重要性。
蛙類的存在還能影響其他生物的行為和分佈。例如,某些蛇類和鳥類的活動模式和棲息地選擇會受到蛙類分佈的影響。在一些濕地生態系統中,蛙類的鳴叫活動不僅吸引了配偶,還為其他依賴聲音定位的動物提供了重要的環境線索。
此外,蛙類在科學研究和環境監測中也扮演著重要角色。由於其對環境變化的敏感性,蛙類常被用作環境毒理學研究的模式生物。通過研究蛙類對特定污染物的反應,科學家們可以評估這些物質對更廣泛生態系統的潛在影響。例如,青蛙胚胎被廣泛用於評估水體中內分泌干擾物的存在和影響。
蛙類的生態角色還體現在它們對微生物群落的影響上。蛙類皮膚上的微生物群落不僅對蛙類自身的健康至關重要,還可能影響周圍環境的微生物平衡。一些研究表明,某些蛙類皮膚上的共生細菌可以抑制有害病原體的生長,這種作用可能延伸到整個生態系統,影響更廣泛的微生物動態。
1.5 蛙類在人類文化中的地位
蛙類在人類文化中占有獨特而重要的地位,其形象和象徵意義貫穿了人類歷史的各個時期和地域。從古老的神話傳說到現代流行文化,蛙類一直是人類想像力和文化表達的重要元素。
在許多古代文明中,蛙類常被賦予神聖或神秘的地位。例如,在古埃及文化中,蛙神黑卡特(Heqet)是生育與重生的象徵。這位女神通常被描繪成蛙頭人身的形象,與尼羅河的泛濫和生命的誕生密切相關。埃及人相信,蛙類的出現預示著豐收和繁榮。在古代美索不達米亞文化中,蛙類也與生育和豐產聯繫在一起,常被視為女神伊什塔爾(Ishtar)的化身。
中國文化中,蛙類同樣具有深厚的文化內涵。「蛙」的諧音與「發」相近,因此常被視為財富和好運的象徵。特別是三腳金蟾的形象,在民間被廣泛認為能招財進寶。這種信仰反映在許多傳統工藝品和裝飾物上,如玉蟾和銅錢蟾等。此外,中國古代神話中的嫦娥奔月故事中,嫦娥在月宮中被變成蟾蜍,這個情節將蛙類與月亮、不死和長生的概念聯繫在一起。
在日本文化中,蛙類也佔有重要地位。日語中「蛙」(カエル, kaeru)與「返る」(かえる, kaeru,意為返回)諧音,因此蛙類常被視為平安歸來的象徵。在日本民間藝術中,常可見到蛙類的形象,特別是在繪畫和雕刻作品中。著名的浮世繪藝術家歌川廣重就創作了許多以蛙為主題的作品,展現了蛙類在日本美學中的獨特地位。
西方文化中,蛙類的形象則更為複雜多樣。在基督教傳統中,蛙類常與瘟疫和災難聯繫在一起,如《聖經》中描述的埃及十災之一就包括蛙災。然而,在歐洲民間傳說中,蛙類又常被賦予正面的魔法力量。最著名的莫過於「青蛙王子」的童話故事,其中被施法變成青蛙的王子通過真愛的吻而恢復人形,這個故事廣為流傳,成為西方文化中變形和救贖主題的經典代表。
在許多原住民文化中,蛙類常被視為雨水和豐收的使者。例如,在澳大利亞原住民的夢幻時期(Dreamtime)傳說中,巨型青蛙提朗巴納(Tiddalik)吞下了世界上所有的水,導致大旱。最終,其他動物設法讓提朗巴納大笑,釋放出水來,恢復了世界的平衡。這個故事不僅反映了蛙類與水的緊密聯繫,也體現了原住民文化中對生態平衡的深刻理解。
在現代流行文化中,蛙類的形象依然廣受歡迎。例如,吉姆·漢森(Jim Henson)創造的科米蛙(Kermit the Frog)成為了全球知名的文化偶像。這個角色不僅在兒童教育節目中發揮了重要作用,還成為了環保運動的代言人,體現了蛙類在現代社會中的積極形象。
文學作品中的蛙類形象也值得關注。在馬克·吐溫(Mark Twain)的短篇小說《著名的跳蛙》(The Celebrated Jumping Frog of Calaveras County)中,一隻會跳遠的青蛙成為了賭博和詐欺的中心,諷刺了人性的貪婪和愚昧。日本作家大江健三郎的小說《飼養蛙》則通過描述一位父親照顧智障兒子的經歷,將蛙類與人性、親情等主題聯繫在一起。
在藝術領域,蛙類也是常見的創作主題。從莫內的睡蓮系列畫作中的青蛙,到現代藝術家如大衛·霍克尼(David Hockney)的青蛙主題作品,蛙類的形象一直吸引著藝術家的創作靈感。這些作品不僅展現了蛙類的生態美學,還常常寓含了藝術家對生命、自然和環境的思考。
在一些文化中,蛙類還與醫療和藥用傳統有關。中國傳統醫學中,蟾酥被認為具有解毒、消腫的功效。在南美洲,某些原住民群體利用箭毒蛙的毒素製作狩獵武器,這種做法不僅反映了人類對自然資源的利用,也展示了蛙類在傳統文化實踐中的重要性。
近年來,隨著環境意識的提升,蛙類在公眾教育和環保運動中的角色日益突出。許多環保組織利用蛙類作為宣傳大使,強調兩棲動物保護的重要性。例如,「拯救青蛙」(Save the Frogs)等組織通過舉辦各種活動,如青蛙日(Frog Day),來提高公眾對兩棲動物保護的認識。
在某些文化中,蛙類還與天氣預報有關。許多農村地區的人們相信,蛙鳴可以預示天氣的變化。這種民間智慧雖然缺乏科學依據,但反映了人類長期以來對自然現象的觀察和解讀。
蛙類在各種儀式和祭祀活動中也扮演著角色。在某些非洲文化中,蛙類被視為雨神的使者,常在祈雨儀式中被提及或使用。這種做法體現了蛙類在人類精神生活和信仰系統中的地位。
現代科技和網絡文化中,蛙類的形象也得到了新的詮釋。例如,網絡表情包中常見的「佩佩蛙」(Pepe the Frog)成為了一種流行文化符號,雖然後來因為被誤用而產生爭議,但其廣泛傳播反映了蛙類形象在當代文化中的適應性和影響力。
2. 蛙類演化與多樣性
蛙類的演化歷程和多樣性是生物學研究中極為引人入勝的主題。這群動物的祖先可追溯至距今約2.65億年前的二疊紀晚期,從最早的四足動物中分化而來。蛙類的演化歷程見證了從完全水生到適應陸地生活的巨大轉變,這個過程塑造了蛙類獨特的形態和生理特徵。
蛙類的多樣性令人驚嘆,這種多樣性不僅體現在物種數量上,更反映在形態、生理和行為的巨大差異中。從體型微小如指甲大小的巴西金蛙(Brachycephalus didactylus),到體長可達32厘米的非洲巨蛙(Conraua goliath),蛙類的體型跨度驚人。
蛙類的棲息環境也極為多樣,從熱帶雨林到乾旱沙漠,從高山到平原,甚至包括一些極端環境。例如,木蛙屬(Rana sylvatica)的某些物種能夠在零下度的環境中存活,通過產生類似防凍劑的物質來保護體內組織。這種適應性反映了蛙類在演化過程中獲得的驚人能力。
蛙類的繁殖策略也展現了豐富的多樣性,從最常見的水中產卵到一些特殊的方式,如背部孵卵、口腔孵卵等。這種多樣性不僅反映了蛙類對不同環境的適應,也為我們理解生殖策略的演化提供了寶貴的案例。
2.1 蛙類演化歷史
蛙類的演化歷史是一段漫長而引人入勝的旅程,揭示了這群獨特動物如何從古老的四足動物祖先演變成今天我們所熟知的形態。這個過程涵蓋了約2.65億年的時間跨度,經歷了多次重大的地質和氣候變遷。
蛙類的祖先可以追溯到二疊紀晚期,當時地球上的生命正經歷著巨大的變革。最早的蛙類化石記錄來自約2.5億年前的馬達加斯加,這個被稱為特里亞索巴特拉庫斯(Triadobatrachus)的物種被認為是現代蛙類的直接祖先。特里亞索巴特拉庫斯已經顯示出一些蛙類的特徵,如延長的後肢和縮短的軀幹,但它仍保留著一些原始特徵,如較長的尾巴。
在三疊紀早期,約2.4億年前,出現了更接近現代蛙類的化石記錄。這些早期蛙類,如原始蛙(Prosalirus bitis),已經具備了跳躍能力,這是蛙類演化史上的一個重要里程碑。跳躍能力的出現不僅改變了蛙類的運動方式,也影響了它們的捕食策略和逃避捕食者的能力。
侏羅紀時期,約2億年前,蛙類開始呈現出更多樣化的趨勢。這個時期出現的諾托巴特拉庫斯(Notobatrachus degiustoi)化石顯示,蛙類已經發展出了類似現代蛙類的骨骼結構。這個時期也見證了蛙類和蟾蜍的分化,形成了兩個主要的演化支系。
白堊紀時期,約1.45億年前,現代蛙類家族開始形成。這個時期的化石記錄顯示,蛙類已經適應了多種生態環境,包括樹棲和地棲的生活方式。例如,來自巴西的化石記錄顯示,早在白堊紀,已經出現了類似現代樹蛙的物種。
蛙類的大規模輻射適應發生在白堊紀末期到第三紀初期,約6600萬年前。這個時期coincided with the extinction of the dinosaurs恐龍滅絕事件,為蛙類的擴張提供了機會。隨著新的生態位的出現,蛙類迅速適應並佔據了各種環境,從熱帶雨林到溫帶草原,甚至包括一些極端環境。
蛙類的演化歷史中一個重要的轉折點是變態過程的出現。從完全水生的生活方式轉變為兼具水陸生活的能力,這種變化不僅涉及形態上的改變,還包括生理和行為的適應。變態過程的演化使蛙類能夠利用不同的生態位,這極大地促進了它們的多樣化。
蛙類的聲音通訊系統也是其演化史上的一個重要創新。雄性蛙類發展出了複雜的發聲器官和行為,用於吸引雌性和劃定領地。這種通訊方式的演化不僅影響了蛙類的繁殖策略,也成為了物種隔離和形成的重要機制。
蛙類的皮膚結構演化也值得關注。與其他陸生脊椎動物不同,蛙類保留了高度通透的皮膚,這使它們能夠通過皮膚進行呼吸和水分交換。這種特性雖然使蛙類更容易受到環境變化的影響,但也使它們能夠適應各種濕潤環境。
在蛙類的演化歷程中,毒性的發展是一個有趣的方面。某些蛙類,特別是新熱帶區的箭毒蛙,演化出了複雜的毒素系統。這些毒素不僅作為防禦機制,還在某些情況下被用作捕食工具。毒性的演化反映了蛙類與捕食者和獵物之間的協同演化關係。
蛙類的繁殖策略也經歷了顯著的演化。從最原始的水中產卵到各種特化的繁殖方式,如背部孵卵、口腔孵卵等,蛙類展現了驚人的適應性。例如,智利袋蟾(Rhinoderma darwinii)演化出了獨特的口腔孵卵方式,雄性將受精卵吞入特化的聲囊中孵化,這種策略在兩棲動物中極為罕見。
氣候變化在蛙類的演化歷史中扮演了重要角色。冰河時期和間冰期的交替導致了蛙類分佈範圍的擴張和收縮,促進了物種的形成和滅絕。例如,歐洲的綠蛙複合體(Pelophylax esculentus complex)的形成就與冰河時期的氣候變化密切相關。
蛙類的形態多樣性也是其演化史的一個重要特徵。從體型微小如指甲大小的巴西金蛙(Brachycephalus didactylus),到體長可達32mm的非洲巨蛙(Conraua goliath),蛙類的體型跨度驚人。這種多樣性反映了蛙類對不同生態位的適應。
蛙類的演化還體現在其生理適應能力上。某些蛙類演化出了極端環境下的生存策略。例如,木蛙屬(Rana sylvatica)的某些物種能夠在零下環境中存活,通過產生類似防凍劑的物質來保護體內組織。這種適應性使蛙類能夠colonize各種極端環境。
近年來,分子生物學技術的發展為我們理解蛙類的演化歷史提供了新的視角。通過對不同蛙類物種的基因序列分析,科學家們能夠重建更詳細的演化樹,揭示以前未知的親緣關係。這些研究不僅幫助我們理解蛙類的演化過程,還為生物地理學研究提供了重要線索。
蛙類的演化歷史還與大陸漂移理論密切相關。通過比較不同大陸上蛙類的親緣關係,科學家們獲得了支持板塊構造理論的重要證據。例如,非洲和南美洲的某些蛙類之間的親緣關係,反映了這兩個大陸曾經相連的歷史。
2.2 蛙類全球多樣性模式
蛙類的全球多樣性模式是生物地理學和生態學研究中一個引人入勝的主題。這種模式不僅反映了蛙類的演化歷史,也體現了地球上不同區域的環境特徵和生態條件。蛙類的分布範圍幾乎涵蓋了除南極洲外的所有大陸,但其多樣性在不同地區呈現出顯著的差異。
熱帶地區,特別是熱帶雨林,是蛙類多樣性的熱點。這些地區不僅擁有最多的蛙類物種,也孕育了許多獨特和稀有的種類。例如,亞馬遜雨林是全球蛙類多樣性最高的地區之一,擁有超過1000種已知的蛙類。這裡不僅物種數量多,而且許多物種都具有獨特的適應特徵。如亞馬遜毒箭蛙(Dendrobates)屬的物種,它們不僅有鮮艷的警戒色,還進化出了強烈的毒性作為防禦機制。
非洲的剛果盆地和東非大裂谷地區也是蛙類多樣性的重要中心。這些地區複雜的地形和多樣的微環境為蛙類的多樣化提供了理想條件。例如,在剛果民主共和國的維龍加國家公園(Virunga National Park),科學家們發現了多種新的蛙類物種,其中包括一些體型極小的物種,反映了這個地區豐富的生態位。
東南亞,尤其是印度尼西亞的蘇門答臘島和婆羅洲,以及菲律賓群島,也是蛙類多樣性的重要地區。這些島嶼的隔離特性促進了物種的特化和演化。例如,在菲律賓的棉蘭老島(Mindanao),科學家們發現了一種能夠在地下生活的蛙類--紫色蛙(Nasikabatrachus puruwaensis),這種蛙類的發現不僅增加了我們對蛙類多樣性的認識,也為理解兩棲動物的演化提供了新的線索。
在溫帶地區,蛙類的多樣性雖然不如熱帶地區豐富,但也呈現出獨特的模式。北美和歐洲的溫帶森林和濕地是許多特有蛙類的家園。例如,北美的樹蛙屬(Hyla)在這些地區呈現出豐富的多樣性,適應了各種不同的生態環境。在歐洲,綠蛙複合體(Pelophylax esculentus complex)的存在展示了蛙類如何通過雜交和多倍體化來適應不同的環境條件。
高山地區雖然環境嚴酷,但也孕育了一些獨特的蛙類物種。例如,在安第斯山脈,科學家們發現了能夠在高海拔地區生存的蛙類,如安第斯蛙屬(Telmatobius)的物種。這些蛙類進化出了特殊的生理機制來應對低溫和低氧環境,展示了蛙類驚人的適應能力。
島嶼生態系統在蛙類的全球多樣性模式中扮演著特殊的角色。由於地理隔離,島嶼上的蛙類常常呈現出高度的特有性。馬達加斯加島就是一個典型的例子,這裡有超過300種特有的蛙類,其中曼蒂拉蛙屬(Mantella)的物種以其鮮艷的色彩和特殊的生態位而聞名。夏威夷群島雖然蛙類物種數量不多,但其唯一的原生蛙類--夏威夷樹蛙(Psychedelic Rock Gecko)的存在,反映了遠距離散布在蛙類分布中的重要性。
在乾旱和半乾旱地區,蛙類雖然種類較少,但卻進化出了一些極為獨特的適應策略。澳大利亞的水盾蛙(Cyclorana platycephala)就是一個很好的例子。這種蛙類能夠在乾旱季節鑽入地下,並分泌一種黏液形成保護層,可以在地下休眠長達數年之久,直到雨季來臨。
溫帶和亞熱帶的季風氣候區域也孕育了豐富的蛙類多樣性。以中國為例,這裡的蛙類多樣性模式反映了複雜的地形和氣候條件。從南方的熱帶雨林到北方的溫帶森林,中國擁有多樣的蛙類fauna,其中包括許多特有種,如峨眉蛙(Rana omeimontis)和秦嶺細痣蛙(Nanorana qinlingensis)。
蛙類的全球多樣性模式還受到歷史地質事件的影響。例如,北美和歐亞大陸的蛙類fauna之間的差異,部分可以歸因於這些大陸在地質歷史上的分離和連接。同樣,南美和非洲蛙類之間的某些相似性,也可以追溯到這兩個大陸曾經相連的歷史。
人類活動對蛙類的全球多樣性模式產生了顯著影響。棲息地破壞、環境污染、氣候變化等因素正在改變蛙類的分布格局。例如,在中美洲,金蛙(Atelopus zeteki)等物種的數量急劇下降,主要是由於真菌性疾病的爆發,這種疾病的傳播可能與全球變暖有關。
另一方面,人為引入的外來種也在改變蛙類的全球分布模式。例如,牛蛙(Lithobates catesbeianus)被引入世界多個地區,對當地的原生蛙類造成了嚴重威脅。這種情況在島嶼生態系統中尤為明顯,如夏威夷群島的原生蛙類就面臨著來自引入種的巨大壓力。
蛙類的全球多樣性模式還與其他生物群體的分布模式存在有趣的關聯。例如,某些地區的蛙類多樣性與植物多樣性呈現正相關,這反映了生態系統複雜性對蛙類多樣性的影響。同時,蛙類的分布模式也影響著其他生物群體,如某些依賴蛙類為食的蛇類或鳥類的分布。
2.3 蛙類適應性輻射
蛙類的適應性輻射是演化生物學中一個引人入勝的主題,展示了這群動物如何在短時間內迅速適應並佔據各種生態位。適應性輻射指的是一個祖先物種在相對較短的時間內,演化出多個具有不同形態、生理和行為特徵的後代物種,以適應不同的環境和生態位。蛙類的適應性輻射為我們提供了理解生物多樣性形成機制的絕佳案例。
馬達加斯加島是研究蛙類適應性輻射的理想地點。這個與非洲大陸分離已久的島嶼孕育了豐富而獨特的蛙類fauna。曼蒂拉蛙屬(Mantella)是一個典型的例子,這個屬的物種展現了驚人的色彩多樣性和生態適應。從生活在雨林地面的金曼蒂拉蛙(Mantella aurantiaca)到棲息在岩石縫隙中的彩虹曼蒂拉蛙(Mantella baroni),每個物種都適應了特定的微環境。這種適應性輻射不僅體現在外觀上,還反映在它們的行為和生理特徵上,如毒性的變化和繁殖策略的多樣化。
中美洲和南美洲的箭毒蛙科(Dendrobatidae)提供了另一個精彩的適應性輻射例子。這個科的成員以其鮮艷的警戒色和強烈的毒性而聞名。然而,不同物種的顏色模式和毒性強度各不相同,反映了它們對不同捕食壓力的適應。例如,草莓毒箭蛙(Oophaga pumilio)在巴拿馬的博卡斯德爾托羅群島(Bocas del Toro)上呈現出驚人的色彩變異,每個島嶼上的種群都有獨特的顏色模式。這種快速的顏色分化被認為是性選擇和天敵壓力共同作用的結果,展示了適應性輻射如何在微觀尺度上進行。
夏威夷群島的樹蛙屬(Eleutherodactylus)提供了一個快速適應性輻射的現代案例。雖然這些蛙類是由人類引入的,但它們在短短幾十年內就適應了各種不同的棲息地,從海岸到山地森林。不同種群開始顯示出形態和行為上的差異,如體型大小、叫聲頻率等的變化,這反映了適應性輻射的初期階段。
菲律賓群島的Platymantis屬蛙類展示了如何島嶼環境可以推動適應性輻射。這個屬的物種在不同島嶼上演化出了各種特化的生活方式,從樹棲到地棲,甚至包括一些特殊的繁殖策略,如直接發育(跳過蝌蚪階段)。這種多樣化反映了蛙類如何利用島嶼環境中的各種生態位。
安第斯山脈的Telmatobius屬蛙類提供了一個高海拔適應性輻射的例子。這個屬的不同物種適應了從山麓到高原湖泊的各種環境。有些物種,如的的喀喀湖蛙(Telmatobius culeus),進化出了特殊的皮膚結構以增加水下呼吸效率,而其他物種則發展出了抵抗高紫外線輻射的能力。這種適應性輻射展示了蛙類如何征服極端環境。
非洲大裂谷地區的蛙類也展現了顯著的適應性輻射。例如,Ptychadena屬的物種在不同的水體環境中特異化,從季節性水坑到永久性湖泊。這些物種在腿部肌肉結構、皮膚濕度調節能力等方面呈現出差異,反映了它們對不同水文環境的適應。
澳大利亞的蛙類提供了乾旱環境適應性輻射的案例。比如,掘穴蛙屬(Neobatrachus)的物種進化出了各種策略來應對乾旱,包括快速發育的蝌蚪、在地下深處休眠的能力等。這種適應性輻射使得蛙類能夠在看似不適合兩棲動物生存的環境中繁衍。
東南亞的峨眉蛙屬(Rana)展示了如何一個廣泛分布的屬可以通過適應性輻射來佔據各種生態位。從高山溪流到低地水田,峨眉蛙屬的不同物種適應了各種水體環境,在體型、皮膤性和繁殖行為等方面呈現出明顯差異。
蛙類的適應性輻射還體現在它們的發聲系統上。不同物種的叫聲頻率、音調和模式各異,這不僅是物種識別的重要特徵,也反映了它們對不同聲學環境的適應。例如,生活在流水環境中的蛙類通常有較高頻率的叫聲,以便在背景噪音中傳播。
蛙類的繁殖策略適應性輻射也十分引人注目。從最常見的水中產卵到一些極為特殊的方式,如背部孵卵、口腔孵卵等,蛙類展現了驚人的創新能力。例如,智利袋蟾(Rhinoderma darwinii)雄性將受精卵存放在特化的聲囊中孵化,這種獨特的繁殖方式是對特定環境壓力的適應結果。
蛙類的適應性輻射還涉及到它們的防禦機制。除了熟為人知的毒性防禦,一些蛙類還進化出了擬態能力。例如,亞馬遜雨林中的某些樹蛙能夠改變體色以mimicking樹皮或葉子,這種適應性不僅幫助它們避免天敵,也提高了捕食效率。
值得注意的是,蛙類的適應性輻射並非總是導致形態上的顯著差異。有時,看似相似的物種可能在生理或行為層面上有重要區別。這種隱蔽的多樣性(cryptic 岐異度)提醒我們,適應性輻射的複雜性遠超過我們的直觀認知。
2.4 全球最奇特的蛙類介紹
透明蛙(Glass frog)。這種生活在中南美洲熱帶雨林的蛙類,其最顯著的特徵是半透明的身體。透過它們的腹部,我們可以清晰地看到內部器官,包括跳動的心臟和消化系統。這種透明性不僅為科學研究提供了便利,也是它們的一種保護色,能夠在樹葉上完美地偽裝自己。
紫色蛙(Purple
frog),這種蛙類生活在印度西高止山脈,因其獨特的紫色皮膚和圓潤的身形而得名。它們的外形與典型的蛙類大不相同,更像是一個小型的馬鈴薯。紫色蛙大部分時間都生活在地下,只有在雨季繁殖時才會短暫出現在地面上。這種生活方式使得它們成為科學家們研究的熱點,也讓它們成為了印度生物多樣性保護的重要對象。
毛蛙(Hairy frog),在南美洲的亞馬遜雨林中,我們可以發現另一種奇特的蛙類。顧名思義,這種蛙的腿部和腰側長有一些類似毛髮的皮膚突起。更令人驚訝的是,當遇到危險時,毛蛙會主動折斷自己腳趾的骨頭,使其穿透皮膚形成利爪,用以防禦敵人。這種極端的自我防禦機制在動物界中非常罕見。
箭毒蛙(Poison dart frog),在中美洲的熱帶雨林中的小型蛙類。它們以鮮豔的體色而聞名,通常是明亮的黃色、藍色或紅色。這些耀眼的顏色實際上是一種警告信號,告訴捕食者它們體內含有劇毒。箭毒蛙的毒素來源於它們的食物,主要是某些特定的螞蟻和跳蟲。有趣的是,失去這些食物來源的箭毒蛙會逐漸失去毒性。
沙漠雨蛙(Desert
rain frog)。這種小巧的蛙類生活在非洲西南部的納米比亞沙漠沿岸地區。它們適應了極度乾燥的環境,主要依靠晨霧和夜間的濕氣來維持水分。沙漠雨蛙的叫聲非常特別,聽起來像是一隻小狗的吠叫,這種聲音在寂靜的沙漠夜晚顯得格外引人注目。
皺褶角蛙(Horned frog)。這種蛙類生活在南美洲的熱帶雨林中,以其巨大的口和獨特的外觀而聞名。它們的頭部有骨質突起,看起來像是長了角,因此得名。皺褶角蛙是典型的伏擊獵手,它們會將自己埋在落葉中,只露出眼睛,靜待獵物經過。當獵物接近時,它們會突然張開大口,一口吞下獵物。這種蛙類能夠吞食比自己體型還大的獵物,包括其他蛙類、蜥蜴,甚至小型哺乳動物。
鼻蛙(Turtle frog),這種蛙類生活在澳大利亞西部的沙漠地區。它們的身體呈圓形,四肢短小,看起來更像一隻小烏龜而非蛙類。鼻蛙最特別的地方在於它們的繁殖方式。與大多數蛙類不同,鼻蛙不需要水域來產卵和孵化蝌蚪。相反,它們會直接在沙土中產下少量的大型卵,幼蛙在卵中完成發育後直接孵化而出。這種獨特的繁殖策略使得鼻蛙能夠在乾旱的環境中生存。
飛蛙(Flying frog),儘管它們並不能真正飛行,但這些生活在東南亞熱帶雨林的蛙類確實能夠在空中滑翔。飛蛙的腳趾間有發達的蹼,當它們從高處跳下時,會將四肢張開,利用這些蹼作為滑翔翼,可以滑翔相當長的距離。這種能力使得飛蛙能夠輕鬆地在樹冠之間移動,躲避地面上的捕食者。
水牛蛙(Smoky
jungle frog)大型蛙類,它們生活在中南美洲的熱帶雨林中,是世界上最大的蛙類之一,體長可達20厘米。水牛蛙因其強壯的後腿和驚人的跳躍能力而聞名,它們能夠輕鬆跳過兩米高的障礙物。這種蛙類的叫聲非常響亮,聽起來像是牛的哞叫,因此得名。水牛蛙是兇猛的捕食者,它們不僅捕食昆蟲和小型脊椎動物,甚至會捕食其他蛙類。
彩虹蛙(Ornate
horned frog),這種蛙類生活在南美洲的熱帶地區,以其鮮豔的體色而聞名。彩虹蛙的背部呈現出綠色、黃色和褐色的複雜花紋,看起來就像一片落葉。這種偽裝不僅能夠幫助它們躲避捕食者,還能夠在獵捕時不被獵物發現。彩虹蛙的嘴巴異常大,能夠吞食比自己頭部還大的獵物。
3. 台灣的蛙類
台灣是一個生物多樣性豐富的島嶼,其獨特的地理位置和多樣化的生態環境為蛙類提供了理想的棲息地。台灣的蛙類物種多樣性相當可觀,分布模式也十分有趣。根據目錄內容,台灣的蛙類主要包括蟾蜍科(Bufonidae)、樹蛙科(Rhacophoridae)、赤蛙科(Ranidae)、狹口蛙科(Microhylidae)和叉舌蛙科(Dicroglossidae)等幾個主要科屬。
每個科屬的蛙類都有其獨特的外觀特徵、棲息地選擇和生活習性。例如,樹蛙科的蛙類通常具有吸盤,適應樹棲生活;而狹口蛙科的蛙類則可能更適應地下或落葉層的生活環境。台灣還擁有一些特有種蛙類,這些物種在演化過程中適應了台灣特有的環境條件,成為台灣生態系統中不可或缺的一部分。
除了本土種類,台灣也面臨外來入侵種蛙類的問題。這些外來種可能對本地生態系統造成影響,如競爭資源或引入新的疾病。因此,了解和管理這些外來種對台灣的蛙類保育至關重要。
台灣還有許多熱門的賞蛙地點,這不僅為自然愛好者提供了觀察蛙類的機會,也提高了公眾對蛙類保育的認識。這些地點可能包括濕地、森林邊緣或山區溪流等多樣化的棲息環境,每個地點都可能有其特色蛙種。
3.1 台灣蛙類物種多樣性
台灣是一個生物多樣性豐富的島嶼,其獨特的地理位置、多樣化的地形和氣候條件為蛙類提供了理想的棲息環境。台灣的蛙類物種多樣性相當可觀,反映了島嶼生態系統的複雜性和演化歷史的深遠影響。
台灣的蛙類主要分為五個科:蟾蜍科(Bufonidae)、樹蛙科(Rhacophoridae)、赤蛙科(Ranidae)、狹口蛙科(Microhylidae)和叉舌蛙科(Dicroglossidae)。這些科屬的存在展示了台灣蛙類的多樣性,每個科都有其獨特的特徵和生態位。
蟾蜍科的蛙類通常體型較大,皮膚粗糙多疣,適應陸地生活。台灣常見的蟾蜍包括盤古蟾蜍和黑眶蟾蜍,這些物種常見於低海拔的平原和丘陵地區,對人為干擾有較強的耐受性。
樹蛙科是台灣最為多樣化的蛙類群體之一,包括諸如莫氏樹蛙、面天樹蛙和台北樹蛙等物種。這些蛙類通常具有吸盤狀的指趾末端,使它們能夠在樹木和其他垂直表面上攀爬。樹蛙的多樣性反映了台灣豐富的森林生態系統,從低地到高山都有不同種類的樹蛙分布。
赤蛙科在台灣同樣具有豐富的多樣性,包括如古氏赤蛙、拉都希氏赤蛙等物種。這些蛙類通常與水域環境緊密相連,常見於溪流、池塘和水田等棲息地。赤蛙科的多樣性反映了台灣豐富的水域生態系統。
狹口蛙科在台灣的代表如小雨蛙和史丹吉氏小雨蛙,這些蛙類體型較小,常棲息於地下或落葉層中。它們的存在豐富了台灣地表下的生物多樣性,在生態系統中扮演著獨特的角色。
叉舌蛙科在台灣的代表包括福建大頭蛙等物種。這些蛙類通常與水域環境關係密切,其多樣性反映了台灣豐富的淡水生態系統。
台灣蛙類的物種多樣性不僅體現在科屬的多樣性上,更體現在各個物種對不同生態位的適應上。例如,在高海拔地區,我們可以發現如褐樹蛙這樣的物種,它們已經適應了寒冷的山區環境。而在低海拔的濕地中,如台北赤蛙這樣的物種則適應了較為溫暖潮濕的環境。
台灣蛙類的多樣性還體現在其特有種的豐富度上。台灣由於長期的地理隔離,形成了許多特有種蛙類,如台北樹蛙、梭德氏赤蛙等。這些特有種的存在不僅增加了台灣的生物多樣性,也為研究島嶼生物地理學和演化生物學提供了寶貴的素材。
台灣蛙類的多樣性還表現在其形態和行為的變異上。例如,在繁殖季節,許多蛙類會呈現出鮮艷的體色,如台北樹蛙雄性在繁殖期會變成亮麗的綠色。而在行為上,不同種類的蛙類也有不同的繁殖策略和鳴叫方式,這些都豐富了台灣的生物多樣性。
然而,台灣的蛙類多樣性也面臨著諸多威脅。棲息地破壞、環境污染、氣候變遷等因素都對台灣的蛙類造成了不同程度的影響。例如,都市化導致的濕地減少直接威脅了許多依賴水域環境的蛙類物種。此外,外來入侵種如斑腿樹蛙的引入也對本地蛙類構成了競爭壓力。
為了保護台灣豐富的蛙類多樣性,科研人員和保育工作者進行了大量的工作。這包括對蛙類物種的調查和監測、棲息地的保護和恢復、以及公眾教育等方面。例如,在墾丁國家公園等保護區內,定期進行的蛙類調查為了解蛙類多樣性的變化提供了重要數據。
台灣蛙類的物種多樣性不僅具有生態學價值,還具有重要的文化和經濟意義。在民間文化中,蛙類常被視為農作物豐收的象徵,反映了人們對自然的敬畏和依賴。在生態旅遊方面,賞蛙活動已成為台灣特色生態旅遊項目之一,為當地社區帶來了經濟效益,同時也提高了公眾對蛙類保育的認識。
台灣蛙類的物種多樣性是一個動態的系統,隨著環境的變化和人類活動的影響而不斷變化。近年來,科研人員通過分子生物學等現代技術,不斷更新對台灣蛙類分類的認識。例如,原本被認為是單一物種的焦眉蝌蚪螈,通過遺傳學研究被確認為包含多個隱藏種,這進一步豐富了我們對台灣蛙類多樣性的認識。
3.2 台灣蛙類分布模式
台灣蛙類的分布模式是一個複雜而有趣的課題,反映了島嶼的地理特徵、氣候條件和生態環境的多樣性。台灣由於其獨特的地理位置和多變的地形,為不同種類的蛙類提供了豐富多樣的棲息環境,從而形成了獨特的分布模式。
台灣的蛙類分布大致可以按照海拔高度、棲息地類型和地理區域來劃分。首先,從海拔角度來看,台灣的蛙類分布呈現出明顯的垂直分層現象。在低海拔地區,如平原和丘陵地帶,常見的蛙類包括澤蛙、虎皮蛙和黑眶蟾蜍等。這些物種適應了較為溫暖潮濕的環境,常在水田、溪流和池塘等水域附近活動。
隨著海拔的升高,蛙類的種類組成也隨之改變。在中海拔山區,如台灣中部的阿里山地區,可以發現如莫氏樹蛙、面天樹蛙等樹棲性強的物種。這些蛙類適應了山區的森林環境,常在樹上或灌木叢中活動。再往上到高海拔地區,如玉山、雪山等地,則可以發現如褐樹蛙、梭德氏赤蛙等耐寒物種。這些高山蛙類已經適應了寒冷的環境,能夠在較低溫度下生存繁衍。
從棲息地類型來看,台灣蛙類的分布也呈現出明顯的差異。水域環境是許多蛙類的重要棲息地,不同類型的水域孕育了不同的蛙類群落。例如,在靜水環境如池塘和水田中,常見的物種包括澤蛙和中國樹蟾。而在流水環境如溪流和瀑布附近,則更容易發現諸如斯文豪氏赤蛙和長腳赤蛙等物種。這些物種在長期演化中形成了特定的形態和行為特徵,以適應不同的水文條件。
森林環境是另一類重要的蛙類棲息地。台灣豐富的森林資源為眾多樹蛙科物種提供了理想的生存空間。例如,在低海拔的闊葉林中,常見白頷樹蛙和橙腹樹蛙等物種。而在較高海拔的針葉林和混合林中,則可能發現諸如褐樹蛙和莫氏樹蛙等適應性較強的物種。這些樹蛙通過其特殊的吸盤結構,能夠在垂直的樹幹和葉片上自如活動,充分利用了森林環境的三維空間。
從地理區域的角度來看,台灣的蛙類分布也呈現出一定的區域性特徵。例如,台灣東部由於受到中央山脈的阻隔,氣候條件和西部有所不同,因此蛙類的組成也有所差異。在東部的花蓮和台東地區,可以發現一些西部較為罕見的物種,如史丹吉氏小雨蛙等。而在台灣南部的恆春半島,由於其獨特的熱帶氣候,孕育了一些特有的蛙類群落,如墾丁樹蛙等。
台灣的離島地區,如綠島和蘭嶼,由於長期的地理隔離,也形成了獨特的蛙類分布模式。例如,在蘭嶼發現的角花綠蛙,就是一個僅分布於該島的特有種。這種分布模式不僅反映了地理隔離對物種演化的影響,也為研究島嶼生物地理學提供了寶貴的素材。
值得注意的是,台灣蛙類的分布模式並非靜態不變的,而是受到多種因素的動態影響。氣候變遷是影響蛙類分布的一個重要因素。近年來,隨著全球氣溫的上升,一些原本只分布在低海拔地區的蛙類,如黑眶蟾蜍,已被發現在較高海拔地區出現。這種分布範圍的擴張可能對當地的生態系統造成影響,同時也反映了蛙類對環境變化的適應能力。
人類活動對台灣蛙類的分布模式也產生了深遠的影響。都市化和農業開發導致的棲息地破壞和片段化,直接改變了許多蛙類的分布範圍。例如,原本廣泛分布於西部平原的台北赤蛙,由於水田和濕地的減少,其分布範圍已大幅縮小。另一方面,人為引入的外來種,如斑腿樹蛙,則擴大了其分布範圍,對本地物種造成競爭壓力。
台灣蛙類的分布模式研究對於生態保育具有重要意義。通過了解不同物種的分布特徵和棲息地需求,可以更有針對性地制定保護措施。例如,對於分布範圍狹窄的特有種,如觀霧山椒魚,可以通過建立保護區等方式來保護其棲息地。而對於廣泛分布但數量下降的物種,如台北樹蛙,則可以通過改善都市綠地設計,建造人工濕地等方式來擴大其適宜棲息地。
此外,台灣蛙類的分布模式研究也為生態系統健康監測提供了重要指標。由於蛙類對環境變化敏感,其分布模式的變化可以反映生態系統的整體狀況。例如,某些對水質要求較高的物種,如梭德氏赤蛙,其分布範圍的變化可以作為水環境質量變化的指標。
台灣蛙類分布模式的研究方法也在不斷更新。除了傳統的野外調查外,近年來利用聲音監測、紅外相機等技術,大大提高了對蛙類分布的監測效率。此外,結合地理信息系統(GIS)和生態位模型等先進工具,科研人員能夠更精確地預測蛙類的潛在分布區域,為保育工作提供科學依據。
3.2.1 台灣的蟾蜍Bufonidae
台灣的蟾蜍科(Bufonidae)是當地兩棲動物群落中重要的一員,雖然種類不如其他科屬豐富,但其獨特的生態地位和適應性使其在台灣生態系統中扮演著關鍵角色。在台灣,蟾蜍科主要包括黑眶蟾蜍和盤古蟾蜍兩種常見物種。
黑眶蟾蜍是台灣最為常見的蟾蜍種類,廣泛分布於全島低海拔地區。其外觀特徵十分鮮明,體型較大,背部呈褐色或灰褐色,佈滿大小不一的疣粒,皮膚粗糙。最顯著的特徵是眼睛周圍有一圈黑色的眶,這也是其名稱的由來。黑眶蟾蜍的體長通常在10到15公分之間,雌性比雄性略大。
盤古蟾蜍則相對較為少見,主要分布在台灣中南部的低海拔地區。其外觀與黑眶蟾蜍相似,但體型稍小,背部顏色較淺,通常呈淺褐色或黃褐色。盤古蟾蜍的皮膚上也佈滿疣粒,但相比黑眶蟾蜍更為細小和密集。其眼睛周圍沒有明顯的黑眶,這是區別兩種蟾蜍的重要特徵之一。
在棲息地選擇方面,台灣的蟾蜍展現出極強的適應性。黑眶蟾蜍幾乎可以在任何有適當濕度的環境中生存,從城市公園到郊區農田,甚至是人類住宅附近都能發現其蹤跡。它們特別喜歡在夜間活動,白天則躲在石頭下、樹洞中或其他陰涼潮濕的地方休息。盤古蟾蜍的棲息地選擇相對較為狹窄,更偏好自然度較高的環境,如森林邊緣、農田周圍的灌木叢等。
蟾蜍的繁殖行為十分有趣。在繁殖季節,通常是在春末夏初的雨季,雄性蟾蜍會聚集在水域附近,發出低沉的鳴叫聲吸引雌性。雌性到達後,雄性會緊緊抱住雌性的背部,這種行為被稱為抱對。雌性會在水中產下大量的卵,同時雄性在卵上釋放精子進行受精。蟾蜍的卵通常呈長串狀,纏繞在水生植物或其他水中物體上。
蝌蚪孵化後會在水中生活一段時間,以藻類和有機碎屑為食。隨著發育,蝌蚪逐漸長出四肢,吸收尾巴,最終變態為小蟾蜍。整個變態過程通常需要一到兩個月的時間,具體取決於環境條件。
在生活習性方面,蟾蜍主要是夜行性動物,白天躲藏,夜晚外出覓食。它們的食性廣泛,主要捕食各種昆蟲、蠕蟲和小型節肢動物。蟾蜍有著獨特的捕食方式,它們會靜坐不動,等待獵物接近,然後迅速伸出黏性的舌頭捕獲獵物。這種捕食行為使蟾蜍成為農業害蟲的天敵,在自然害蟲控制中發揮重要作用。
蟾蜍科動物還有一個顯著特徵,就是在體側或頸部有一對大型的耳後腺,俗稱毒腺。這些腺體能分泌一種白色的毒性物質,可以有效地防禦天敵。雖然這種毒素對人類皮膚接觸通常不會造成嚴重傷害,但如果接觸到眼睛或黏膜,可能會引起刺激和不適。
在豐富度方面,黑眶蟾蜍在台灣的數量相當可觀,幾乎在所有適宜的低海拔地區都能發現其蹤跡。這種廣泛分布和高豐富度反映了黑眶蟾蜍極強的環境適應能力和繁殖能力。相比之下,盤古蟾蜍的數量和分布範圍較為有限,主要集中在台灣中南部的特定區域。
蟾蜍在台灣生態系統中扮演著重要角色。作為中間消費者,它們控制昆蟲等小型動物的數量,同時也是蛇類、鳥類等高級消費者的食物來源。此外,由於蟾蜍對環境變化較為敏感,特別是對水質和空氣污染,因此常被用作環境健康的生物指標。
然而,台灣的蟾蜍也面臨著一些威脅。棲息地破壞和片段化是主要問題之一,特別是對分布範圍較窄的盤古蟾蜍影響更大。此外,道路建設導致的路殺也是蟾蜍面臨的重要威脅,特別是在繁殖季節,大量蟾蜍橫穿道路時容易遭到車輛碾壓。
為了保護台灣的蟾蜍,相關部門和保育組織採取了多項措施。例如,在一些重要的蟾蜍棲息地建立保護區,限制人為干擾。在繁殖季節,一些地區會設立臨時的蟾蜍通道或警示牌,減少路殺事件的發生。同時,通過環境教育提高公眾對蟾蜍生態價值的認識,也是保護工作的重要一環。
3.2.2 台灣的樹蛙Rhacophoridae
台灣樹蛙科(Rhacophoridae)是台灣兩棲動物中最為豐富多樣的群體之一,在生態系統中扮演著重要角色。這個科屬的蛙類以其獨特的外觀、巧妙的棲息地選擇和有趣的繁殖行為而聞名。
在台灣,樹蛙科的常見種類包括白頷樹蛙、莫氏樹蛙、面天樹蛙、褐樹蛙和台北樹蛙等。每個物種都有其獨特的特徵和生態位。
白頷樹蛙是台灣最常見的樹蛙之一,廣泛分布於低海拔至中海拔地區。其最顯著的特徵是下巴和喉部的白色斑塊,體色則可能因環境和情緒而變化,從淺綠色到深褐色不等。白頷樹蛙的體型中等,通常體長在3到5公分之間。
莫氏樹蛙是另一種常見的樹蛙,其特徵是背部有明顯的X形花紋,體色通常為褐色或灰褐色,能夠很好地融入樹皮或落葉的背景中。這種保護色使得莫氏樹蛙在靜止時難以被發現,是其避免天敵的重要策略。
面天樹蛙因其獨特的姿勢而得名,它們常常仰頭朝天,彷彿在觀察天空。這種樹蛙體型較小,通常不超過3公分,體色多變,可以根據環境快速改變顏色,從綠色到褐色都有可能。
褐樹蛙是台灣高海拔地區的特色物種,能夠適應較低的溫度。它們的體色以褐色為主,背部常有不規則的深色斑紋。褐樹蛙的體型相對較大,成體可達5到7公分。
台北樹蛙是台灣的特有種,主要分布在北部地區。它們的體色通常為綠色或黃綠色,背部有細小的顆粒狀突起,這些突起在繁殖季節會變得更加明顯。
在棲息地選擇方面,樹蛙科的物種展現出極強的樹棲傾向。它們的腳趾末端都有吸盤狀結構,使其能夠輕鬆地在垂直的樹幹或光滑的葉片上攀爬。白頷樹蛙和莫氏樹蛙常見於低海拔的闊葉林和竹林中,也能適應城市公園和果園等人為環境。面天樹蛙則更偏好濕潤的環境,常在溪流附近的植被上發現。褐樹蛙主要生活在較高海拔的針葉林和混合林中,能夠忍受較低的溫度。台北樹蛙則多見於北部的低海拔森林和農業區。
樹蛙的繁殖行為十分有趣,展現了多樣的繁殖策略。大多數樹蛙種類採用水域繁殖的方式,在繁殖季節,雄性會聚集在水邊或樹上發出響亮的鳴叫聲吸引雌性。例如,白頷樹蛙的鳴叫聲類似「格格格」的重複音,而莫氏樹蛙則發出類似「嘰嘰」的高頻聲音。
一些樹蛙種類還發展出了特殊的繁殖策略。面天樹蛙就有著獨特的泡沫巢繁殖方式。雌性會在樹葉上產下卵泡,同時分泌黏液並用後肢拍打,形成一個泡沫狀的巢。這個泡沫巢不僅能保護卵免受乾燥和捕食,還能為發育中的胚胎提供必要的濕度和氧氣。
褐樹蛙則採用了另一種策略來適應高海拔的寒冷環境。它們會將卵產在潮濕的地面上,而不是直接產在水中。這種陸地繁殖的方式可以避免卵和幼體在寒冷的水中發育過慢的問題。
在生活習性方面,大多數樹蛙都是夜行性動物,白天躲在樹葉或樹洞中休息,傍晚到夜間則活躍起來捕食。它們主要以各種昆蟲為食,如蚊子、蛾類和甲蟲等。樹蛙的舌頭非常靈活,能夠快速彈出捕獲飛行中的昆蟲。
樹蛙在生態系統中扮演著重要角色。作為昆蟲的捕食者,它們有助於控制昆蟲數量,包括一些農業害蟲。同時,樹蛙也是許多鳥類、蛇類和小型哺乳動物的食物來源,是食物鏈中的重要一環。
在豐富度方面,不同種類的樹蛙表現各異。白頷樹蛙和莫氏樹蛙由於適應性強,在台灣的數量相當可觀,幾乎在所有適宜的低中海拔地區都能發現。面天樹蛙雖然分布廣泛,但數量相對較少。褐樹蛙由於棲息地限制,主要集中在特定的高海拔地區,數量相對有限。台北樹蛙作為特有種,其分布範圍較為局限,主要集中在北部地區,數量也相對較少。
然而,台灣的樹蛙也面臨著一些威脅。棲息地破壞和片段化是主要問題之一,特別是對於分布範圍較窄的特有種影響更大。此外,氣候變遷也對樹蛙造成影響,尤其是對高海拔物種如褐樹蛙的影響更為明顯。隨著氣溫升高,這些適應寒冷環境的物種可能面臨棲息地縮小的問題。
為了保護台灣的樹蛙,相關部門和保育組織採取了多項措施。例如,在一些重要的樹蛙棲息地建立保護區,限制人為干擾。同時,通過環境教育提高公眾對樹蛙生態價值的認識,鼓勵民眾參與到樹蛙保育工作中來。一些地區還開展了「賞蛙」活動,既能提高公眾對樹蛙的了解和興趣,又能為當地帶來生態旅遊收入,促進保育工作的開展。
研究人員也在持續進行樹蛙的生態學研究,包括監測樹蛙種群數量變化、研究氣候變遷對樹蛙的影響等。這些研究不僅有助於更好地了解樹蛙的生態需求,也為制定更有效的保育策略提供了科學依據。
3.2.3 台灣的赤蛙Ranidae
台灣的赤蛙科(Ranidae)是當地兩棲動物中具有重要生態地位的一個群體,其種類豐富,生態習性多樣,在台灣的水域和陸地生態系統中扮演著關鍵角色。赤蛙科的蛙類通常與水域環境密切相關,但也有一些物種能夠適應較為乾燥的環境。
在台灣,赤蛙科的常見種類包括梭德氏赤蛙、拉都希氏赤蛙、貢德氏赤蛙、斯文豪氏赤蛙和台北赤蛙等。每個物種都有其獨特的特徵和生態位。
梭德氏赤蛙是台灣中高海拔地區常見的物種,其最顯著的特徵是背部有兩條明顯的背側褶,體色通常為褐色或紅褐色,背部有不規則的深色斑點。這種赤蛙的體型中等,成體體長通常在5到8公分之間。梭德氏赤蛙的另一個特點是其皮膚粗糙,有許多細小的疣粒,這些特徵有助於它們在森林環境中進行保護色偽裝。
拉都希氏赤蛙是台灣低海拔地區常見的物種,體型較小,成體體長通常不超過5公分。它們的體色變化較大,從淺褐色到深褐色都有,背部常有不規則的深色斑紋。拉都希氏赤蛙的特徵之一是其後肢較長,使它們能夠進行長距離的跳躍,這在逃避天敵時非常有利。
貢德氏赤蛙是台灣特有種,主要分布在中低海拔山區。這種赤蛙的體型較大,成體體長可達7到10公分。它們的體色通常為褐色或灰褐色,背部有不規則的深色斑紋。貢德氏赤蛙的特點是其鼓膜特別明顯,雄性在繁殖季節時喉部會變成黑色。
斯文豪氏赤蛙是台灣低海拔地區常見的大型赤蛙,成體體長可達10公分以上。它們的體色通常為綠色或褐綠色,背部有不規則的深色斑點。斯文豪氏赤蛙的特徵是其皮膚光滑,背部有兩條明顯的背側褶。這種赤蛙的後肢特別發達,使它們能夠進行強有力的跳躍。
台北赤蛙是台灣特有種,主要分布在北部低海拔地區。這種赤蛙的體型中等,成體體長約5到7公分。它們的體色通常為褐色或紅褐色,背部有不規則的深色斑紋。台北赤蛙的特點是其皮膚較為光滑,背部的斑紋常呈現出類似樹葉脈絡的形狀,這種特徵有助於它們在落葉中進行偽裝。
在棲息地選擇方面,赤蛙科的物種展現出與水域環境密切相關的特點,但不同種類對棲息地的要求也有所不同。梭德氏赤蛙主要生活在中高海拔的山區溪流附近,它們喜歡在溪流邊的岩石或落葉堆中活動。拉都希氏赤蛙則更為適應人為環境,常見於低海拔的水田、溝渠和池塘附近。貢德氏赤蛙偏好山區的溪流和水潭,常在水邊的石縫或落葉堆中棲息。斯文豪氏赤蛙生活在低海拔的各種水域環境中,包括池塘、水田和緩流的河川。台北赤蛙則主要生活在低海拔的水田和濕地環境中。
赤蛙的繁殖行為通常與水域環境密切相關。在繁殖季節,雄性赤蛙會聚集在水邊或水中發出響亮的鳴叫聲吸引雌性。不同種類的赤蛙有不同的鳴叫聲,這種聲音不僅用於吸引配偶,也用於劃分領域。例如,梭德氏赤蛙的鳴叫聲類似「咯-咯-咯」的重複音,而斯文豪氏赤蛙則發出類似「呱-呱-呱」的低沉叫聲。
赤蛙的產卵通常在水中進行,雌性會一次產下大量的卵,這些卵通常呈球形卵塊,漂浮在水面或附著在水生植物上。例如,斯文豪氏赤蛙的卵塊可以含有數千個卵。卵孵化後,幼蝌蚪會在水中生活一段時間,逐漸發育四肢並吸收尾巴,最終變態為小蛙。
在生活習性方面,大多數赤蛙都是晝夜活動的動物,但在夜間活動更為頻繁。它們主要以各種昆蟲、蠕蟲和小型節肢動物為食。赤蛙的捕食方式通常是靜候獵物接近,然後快速伸出舌頭將獵物捕獲。一些大型的赤蛙,如斯文豪氏赤蛙,甚至能捕食小型魚類和其他蛙類。
赤蛙在生態系統中扮演著重要角色。作為中間消費者,它們控制昆蟲等小型動物的數量,同時也是蛇類、鳥類等高級消費者的食物來源。此外,由於赤蛙對水質變化較為敏感,因此常被用作水環境質量的生物指標。
在豐富度方面,不同種類的赤蛙表現各異。拉都希氏赤蛙和斯文豪氏赤蛙由於適應性強,在台灣低海拔地區的數量相當可觀。梭德氏赤蛙在適宜的中高海拔地區也有較高的豐富度。相比之下,台北赤蛙和貢德氏赤蛙作為特有種,其分布範圍較為局限,數量相對較少,面臨著更大的保育壓力。
然而,台灣的赤蛙也面臨著諸多威脅。棲息地破壞和汙染是主要問題之一,特別是對於依賴特定水域環境的物種影響更大。例如,台北赤蛙的棲息地水田因都市化而大幅減少,導致其數量急劇下降。此外,外來種的入侵也對本地赤蛙造成威脅,如入侵的福建大頭蛙會捕食本地赤蛙的幼體。
為了保護台灣的赤蛙,相關部門和保育組織採取了多項措施。例如,在一些重要的赤蛙棲息地建立保護區,限制人為干擾。同時,通過環境教育提高公眾對赤蛙生態價值的認識,鼓勵民眾參與到赤蛙保育工作中來。一些地區還開展了棲息地復育計畫,如在都市地區建造生態池塘,為赤蛙提供適宜的棲息環境。
3.2.4 台灣的狹口蛙Microhylidae
台灣的狹口蛙科(Microhylidae)是一個獨特而有趣的蛙類群體,雖然種類不如其他科屬豐富,但其特殊的生態位和生活習性使其在台灣的生態系統中佔有重要地位。狹口蛙科的蛙類以其小巧的體型和獨特的形態而聞名,在台灣主要有兩個常見種類:小雨蛙和史丹吉氏小雨蛙。
小雨蛙是台灣最為常見的狹口蛙種類,廣泛分布於全島低海拔地區。其最顯著的特徵是極小的體型,成體體長通常只有2到3公分,是台灣體型最小的蛙類之一。小雨蛙的體型呈橢圓形,頭部較小,吻端鈍圓。它們的皮膚光滑,體色變化較大,從淺褐色到深褐色不等,背部常有不規則的深色斑紋或條紋。小雨蛙的腹部通常呈白色或淺黃色,有時會有細小的斑點。最引人注目的是它們的眼睛,相對於小巧的體型而言顯得格外大而明亮。
史丹吉氏小雨蛙則相對較為少見,主要分布在台灣東部和南部的低海拔地區。其外觀與小雨蛙相似,但體型略大,成體體長可達3到4公分。史丹吉氏小雨蛙的體色通常較為鮮艷,背部常呈現紅褐色或橙褐色,有時會有亮麗的黃色或橙色斑點。它們的腹部通常為白色或淺黃色,有時會有深色的斑點或網紋。
在棲息地選擇方面,狹口蛙科的蛙類展現出獨特的偏好。小雨蛙和史丹吉氏小雨蛙都是地棲性蛙類,喜歡生活在潮濕的環境中。它們常見於落葉堆、腐爛的樹幹下、石頭縫隙或鬆軟的土壤中。這些蛙類特別喜歡在下雨後出來活動,這也是「雨蛙」名稱的由來。
小雨蛙的適應性較強,能夠在多種環境中生存,從自然森林到都市公園都能發現其蹤跡。它們特別喜歡生活在潮濕的花園、苗圃或農田附近。相比之下,史丹吉氏小雨蛙對環境的要求更為嚴格,更偏好保存較好的自然環境,如原始林邊緣或次生林中。
狹口蛙的繁殖行為十分有趣。在繁殖季節,通常是在春末夏初的雨季,雄性狹口蛙會聚集在臨時性的水域附近,如雨後形成的小水窪或積水坑。雄蛙會發出獨特的鳴叫聲吸引雌性。小雨蛙的叫聲類似「咯-咯-咯」的快速重複音,聲音雖然細小但在安靜的夜晚仍能傳播較遠的距離。
雌性到達後,雄性會抱住雌性的腰部,這種行為被稱為腰抱。雌性會在水中產下大量的小型卵粒,通常是成串或成片漂浮在水面上。受精後的卵會迅速發育,通常在24到48小時內就能孵化。狹口蛙的蝌蚪發育速度非常快,這是對其常選擇臨時性水體作為繁殖場所的一種適應。從卵孵化到幼蛙上岸,整個過程可能只需要兩到三週的時間。
在生活習性方面,狹口蛙主要是夜行性動物,白天躲藏在陰涼潮濕的地方,夜晚外出覓食。它們的食性相當特殊,主要以螞蟻和白蟻等小型昆蟲為食。狹口蛙有著特化的口器結構,能夠有效地捕食這些小型獵物。它們的舌頭較短但黏性很強,可以快速彈出捕獲獵物。
狹口蛙在生態系統中扮演著獨特的角色。作為專門的螞蟻和白蟻捕食者,它們有助於控制這些昆蟲的數量,維持生態平衡。同時,由於其小巧的體型,狹口蛙也是許多爬行動物和鳥類的食物來源,在食物鏈中佔有重要地位。
在豐富度方面,小雨蛙在台灣的數量相當可觀,幾乎在所有適宜的低海拔地區都能發現。這種廣泛分布和高豐富度反映了小雨蛙極強的環境適應能力和繁殖能力。相比之下,史丹吉氏小雨蛙的數量和分布範圍較為有限,主要集中在台灣東部和南部的特定區域。
然而,台灣的狹口蛙也面臨著一些威脅。棲息地破壞和環境污染是主要問題之一。由於狹口蛙對環境濕度的高度依賴,乾旱或水源污染都可能對其造成嚴重影響。此外,農藥的過度使用也是一個潛在的威脅,因為這可能減少狹口蛙的食物來源或直接毒害蛙類。
氣候變遷也對狹口蛙構成挑戰。極端氣候事件,如長期乾旱或異常降雨,可能改變狹口蛙的棲息環境,影響其繁殖和生存。特別是對於依賴臨時性水體繁殖的狹口蛙來說,降雨模式的改變可能直接影響其繁殖成功率。
為了保護台灣的狹口蛙,相關部門和保育組織採取了多項措施。例如,在一些重要的狹口蛙棲息地建立保護區,限制人為干擾。同時,通過環境教育提高公眾對狹口蛙生態價值的認識,鼓勵民眾參與到蛙類保育工作中來。
在都市地區,一些社區和學校開始推廣「友善蛙類」的園藝實踐,如減少農藥使用,保留一些自然植被區域,為狹口蛙提供棲息環境。這不僅有助於保護狹口蛙,也為都市生態增添了活力。
研究人員也在持續進行狹口蛙的生態學研究,包括監測種群數量變化、研究氣候變遷對狹口蛙的影響等。這些研究不僅有助於更好地了解狹口蛙的生態需求,也為制定更有效的保育策略提供了科學依據。
3.2.5 台灣的叉舌蛙Dicroglossidae
台灣的叉舌蛙科(Dicroglossidae)是當地兩棲動物中一個獨特而重要的群體,雖然種類不如其他科屬豐富,但其特殊的生態位和生活習性在台灣的生態系統中佔有重要地位。叉舌蛙科的蛙類以其獨特的舌頭結構而得名,在台灣主要有兩個常見種類:福建大頭蛙和海蛙。
福建大頭蛙是台灣最為常見的叉舌蛙種類,廣泛分布於全島低海拔地區。其最顯著的特徵是大而寬的頭部,這也是其名稱的由來。成體體長通常在7到10公分之間,是台灣體型較大的蛙類之一。福建大頭蛙的體型強壯,後肢發達,有利於跳躍和游泳。它們的皮膚較為光滑,體色通常為褐色或灰褐色,背部有不規則的深色斑紋。腹部則呈現淺色,常帶有斑點或網紋。福建大頭蛙的眼睛較大,瞳孔呈水平橢圓形,這是其夜行性生活方式的一個適應。
海蛙則相對較為少見,主要分布在台灣西部和南部的低海拔沿海地區。其外觀與福建大頭蛙有所不同,體型略小,成體體長通常在5到7公分之間。海蛙的體色通常較為鮮艷,背部可能呈現綠色、褐色或灰色,常有明顯的條紋或斑點。它們的皮膚上有許多細小的疣粒,這是其適應鹽水環境的一種特徵。海蛙的後肢特別發達,趾間蹼也較為明顯,這些特徵都有助於其在水中的活動。
在棲息地選擇方面,叉舌蛙科的蛙類展現出對水域環境的強烈偏好。福建大頭蛙是一種半水棲的蛙類,喜歡生活在各種淡水環境中,如水田、池塘、溝渠和緩流的河川等。它們能夠適應多種環境,從自然水域到人工水體都能生存。在非繁殖季節,福建大頭蛙也會離開水域,在附近的草叢或石縫中棲息。
相比之下,海蛙對棲息地的選擇更為特殊。它們主要生活在沿海地區的鹹水或半鹹水環境中,如紅樹林、鹽田、河口濕地等。海蛙能夠忍受一定程度的鹽分,這是其他台灣蛙類所不具備的特性。這種獨特的棲息地選擇使海蛙在生態系統中佔據了特殊的生態位。
叉舌蛙的繁殖行為與其他蛙類相似,但也有其獨特之處。在繁殖季節,通常是在春末夏初的雨季,雄性叉舌蛙會聚集在水域附近,發出響亮的鳴叫聲吸引雌性。福建大頭蛙的叫聲低沉有力,類似「呱-呱-呱」的重複音,在夜晚能傳播較遠的距離。海蛙的叫聲則相對較為輕柔,類似「咕-咕-咕」的快速重複音。
雌性到達後,雄性會抱住雌性的腰部進行交配。雌性通常會在水中產下大量的卵,這些卵呈球形卵塊,漂浮在水面或附著在水生植物上。福建大頭蛙的卵塊特別大,一次可產下數千個卵。卵孵化後,幼蝌蚪會在水中生活一段時間,逐漸發育四肢並吸收尾巴,最終變態為小蛙。整個變態過程通常需要一到兩個月的時間,具體取決於環境條件。
在生活習性方面,叉舌蛙主要是夜行性動物,白天通常躲在水中或潮濕的地方休息,夜晚則活躍起來捕食。它們的食性廣泛,主要捕食各種昆蟲、蠕蟲和小型節肢動物。福建大頭蛙由於體型較大,甚至能捕食小型魚類和其他蛙類。叉舌蛙有著特殊的舌頭結構,舌頭前端分叉,這使它們能夠更有效地捕捉和控制獵物。
叉舌蛙在生態系統中扮演著重要角色。作為中間消費者,它們控制昆蟲等小型動物的數量,同時也是蛇類、鳥類等高級消費者的食物來源。特別是海蛙,作為少數能夠適應鹹水環境的蛙類,在沿海生態系統中佔據了獨特的生態位置。
在豐富度方面,福建大頭蛙在台灣的數量相當可觀,幾乎在所有適宜的低海拔水域環境中都能發現。這種廣泛分布和高豐富度反映了福建大頭蛙極強的環境適應能力和繁殖能力。相比之下,海蛙的數量和分布範圍較為有限,主要集中在台灣西部和南部的特定沿海區域。
然而,台灣的叉舌蛙也面臨著一些威脅。棲息地破壞和水質污染是主要問題之一。隨著都市化和工業化的發展,許多水域環境被填埋或污染,直接影響了叉舌蛙的生存空間。特別是對於海蛙來說,沿海濕地的減少和海岸線的人工化是一大威脅。此外,外來入侵種的競爭也是一個潛在問題,例如入侵的福建大頭蛙(雖然同屬於叉舌蛙科,但並非台灣原生種)可能與本地種競爭資源。
氣候變遷也對叉舌蛙構成挑戰。極端氣候事件,如長期乾旱或異常降雨,可能改變叉舌蛙的棲息環境,影響其繁殖和生存。對於海蛙來說,海平面上升可能導致其棲息地被淹沒或鹽分含量改變,這對其生存造成威脅。
為了保護台灣的叉舌蛙,相關部門和保育組織採取了多項措施。例如,在一些重要的叉舌蛙棲息地建立保護區,限制人為干擾。同時,通過環境教育提高公眾對叉舌蛙生態價值的認識,鼓勵民眾參與到蛙類保育工作中來。對於海蛙,沿海濕地的保護和恢復工作尤為重要。
研究人員也在持續進行叉舌蛙的生態學研究,包括監測種群數量變化、研究氣候變遷對叉舌蛙的影響等。這些研究不僅有助於更好地了解叉舌蛙的生態需求,也為制定更有效的保育策略提供了科學依據。
3.3 台灣蛙類特有種
台灣作為一個獨特的島嶼生態系統,孕育了眾多特有物種,其中蛙類特有種更是台灣生物多樣性的重要組成部分。這些特有種蛙類經過長期的地理隔離和演化,形成了獨特的形態特徵和生態習性,反映了台灣獨特的自然環境和演化歷程。
在台灣已知的蛙類中,特有種的數量相當可觀,約佔台灣蛙類總數的三分之一。這些特有種分布在不同的科屬中,包括樹蛙科、赤蛙科和狹口蛙科等。每一種特有蛙類都有其獨特的生態位和適應特徵,為台灣的生態系統增添了豐富的色彩。
台北樹蛙是台灣最為知名的特有種之一,屬於樹蛙科。這種蛙類主要分布在台灣北部的低海拔山區,體型中等,成體體長約3到4公分。台北樹蛙的外觀十分美麗,體色通常為翠綠色或黃綠色,有時會有細小的金色斑點。它們的皮膚光滑,指趾末端有明顯的吸盤,適合在樹上活動。台北樹蛙的繁殖行為十分有趣,雄蛙會在樹葉上建造泡沫巢,雌蛙將卵產在泡沫巢中,這種獨特的繁殖策略有助於保護卵和幼體免受乾燥和捕食。
梭德氏赤蛙是另一種重要的台灣特有種,屬於赤蛙科。這種蛙類主要分布在台灣中高海拔的山區,適應了較為寒冷的環境。梭德氏赤蛙的體型較大,成體體長可達7到9公分。它們的體色通常為褐色或紅褐色,背部有不規則的深色斑點,皮膚上有許多細小的疣粒。梭德氏赤蛙是優秀的跳躍者,後肢特別發達。它們通常生活在山區溪流附近,繁殖時會在溪流中的石頭縫隙產卵,這種繁殖策略是對山區環境的一種適應。
盤古蟾蜍是台灣特有的蟾蜍科物種,主要分布在台灣中南部的低海拔地區。這種蟾蜍的體型中等,成體體長約6到8公分。盤古蟾蜍的外觀與一般蟾蜍相似,體色為褐色或灰褐色,皮膚粗糙,佈滿疣粒。它們的眼睛周圍有明顯的山脊狀突起,這是其獨特的特徵。盤古蟾蜍主要生活在森林邊緣和農田附近,對人為干擾有一定的耐受性。
史丹吉氏小雨蛙是台灣特有的狹口蛙科物種,主要分布在台灣東部和南部的低海拔地區。這種小型蛙類的體長通常不超過3公分,是台灣體型最小的蛙類之一。史丹吉氏小雨蛙的體色鮮艷,通常為紅褐色或橙褐色,有時會有明亮的黃色或橙色斑點。它們主要生活在潮濕的落葉層或腐爛的樹幹下,繁殖時會選擇臨時性的小水窪。
台灣特有種蛙類的生態習性和適應特徵反映了台灣獨特的自然環境。例如,許多特有種樹蛙發展出了在樹上產卵的策略,這可能是對台灣多雨潮濕氣候的一種適應。而一些特有種赤蛙則適應了台灣山區的環境,能夠在較低溫度下活動和繁殖。
特有種蛙類的分布模式也反映了台灣的地理特徵。一些特有種,如台北樹蛙,主要分布在特定的地理區域,這可能與台灣的地形分割和氣候差異有關。而一些特有種,如梭德氏赤蛙,則廣泛分布在全島的山區,顯示出對山地環境的良好適應。
台灣特有種蛙類面臨的主要威脅包括棲息地破壞、環境污染和氣候變遷。例如,台北樹蛙的棲息地因都市化而不斷縮小,直接威脅到其生存。而山區特有種如梭德氏赤蛙則可能受到氣候變遷的影響,溫度升高可能導致其適宜棲息地向更高海拔移動。
為了保護這些珍貴的特有種蛙類,台灣採取了多項保育措施。例如,設立特有種蛙類的重要棲息地為保護區,限制人為干擾。同時,通過環境教育提高公眾對特有種蛙類的認識和保護意識。一些地區還開展了棲息地復育計畫,如在都市地區建造生態池塘,為特有種蛙類提供繁殖場所。
研究人員也在持續進行特有種蛙類的生態學研究,包括基因多樣性分析、生態位研究和種群動態監測等。這些研究不僅有助於更好地了解特有種蛙類的生態需求和演化歷史,也為制定更有效的保育策略提供了科學依據。
台灣特有種蛙類不僅具有重要的生態價值,也具有獨特的文化和教育意義。它們是台灣自然遺產的重要組成部分,反映了台灣獨特的自然環境和演化歷程。通過保護這些特有種蛙類,我們不僅在維護生態平衡,也在保護台灣的自然文化遺產。
3.4 台灣蛙類外來入侵種與影響
台灣作為一個島嶼生態系統,其獨特的生物多樣性一直備受關注。然而,隨著全球化的進程,外來入侵種對台灣本地生態系統的威脅日益嚴重。在蛙類中,外來入侵種的問題尤為突出,對台灣的生態環境造成了多方面的影響。
在台灣,最為人所知的蛙類外來入侵種是斑腿樹蛙。這種原產於東南亞的樹蛙於1990年代末期被引入台灣,最初可能是作為寵物或實驗動物。斑腿樹蛙體型中等,成體體長可達5至8公分,體色多變,通常為褐色或灰色,腿部有明顯的條紋,這也是其名稱的由來。斑腿樹蛙具有極強的適應能力和繁殖力,能夠在多種環境中生存,包括城市、農田和自然森林。
斑腿樹蛙的入侵對台灣的生態系統造成了多重影響。首先,它們與本地蛙類競爭食物和棲息地資源。斑腿樹蛙的食性廣泛,能夠捕食各種昆蟲、小型無脊椎動物,甚至是其他蛙類的幼體。這種廣泛的食性使得斑腿樹蛙能夠在多種環境中生存,同時也可能導致本地物種的食物資源減少。
其次,斑腿樹蛙可能攜帶對本地蛙類有害的病原體。例如,有研究發現斑腿樹蛙可能攜帶壺菌,這種真菌對許多兩棲動物都有致命的影響。雖然目前在台灣尚未發現由斑腿樹蛙引起的大規模壺菌感染,但這種潛在風險仍然存在。
此外,斑腿樹蛙的大量繁殖也可能改變當地的生態平衡。在繁殖季節,大量的斑腿樹蛙聚集在水域附近,其鳴叫聲可能干擾本地蛙類的繁殖活動。同時,大量的斑腿樹蛙幼體可能與本地蛙類的幼體競爭有限的水域資源。
另一種值得關注的外來入侵蛙類是福建大頭蛙。雖然福建大頭蛙在分類上屬於叉舌蛙科,與台灣的一些本地種類相近,但它並非台灣的原生種。福建大頭蛙體型較大,成體體長可達10公分以上,具有強壯的後肢和大而寬的頭部。這種蛙類主要生活在水域環境中,如水田、池塘和溪流。
福建大頭蛙的入侵同樣對台灣的生態系統造成了影響。由於其體型較大,福建大頭蛙可能捕食本地的小型蛙類和其他水生動物。此外,福建大頭蛙的繁殖力強,一次可產下數千個卵,這使得它們能夠在短時間內快速擴張種群。
福建大頭蛙的入侵還可能影響水域生態系統的平衡。作為一種大型的水棲蛙類,福建大頭蛙可能改變水域環境的營養循環和食物網結構。同時,它們的大量繁殖可能影響水質,特別是在一些小型或封閉的水體中。
面對這些外來入侵蛙類的威脅,台灣的研究人員和保育工作者採取了多種措施。首先,加強對外來入侵種的監測和研究,了解它們的分布範圍、生態習性和對本地生態系統的影響。例如,研究人員利用聲音監測技術來追蹤斑腿樹蛙的分布和活動模式。
其次,開展外來入侵種的移除工作。在一些重要的生態區域,如國家公園和自然保護區,進行有針對性的外來入侵蛙類清除行動。這些行動通常包括手工捕捉成體和清除卵塊等方法。然而,由於外來入侵蛙類的繁殖能力強,這種移除工作需要長期持續進行才能取得效果。
同時,加強對外來寵物貿易的管理也是防止新的外來入侵種引入的重要措施。通過法規限制和公眾教育,減少外來蛙類被引入和釋放到自然環境中的機會。
此外,保護和恢復本地蛙類的棲息地也是應對外來入侵種威脅的重要策略。通過改善本地蛙類的生存環境,增強其競爭力,可以在一定程度上減少外來入侵種的影響。
在公眾教育方面,台灣的保育組織和教育機構也做了大量工作。通過各種形式的宣傳和教育活動,提高公眾對外來入侵蛙類危害的認識,鼓勵公眾參與到外來入侵種的監測和防控工作中來。
儘管面臨諸多挑戰,台灣在應對蛙類外來入侵種方面已經取得了一些成果。例如,在一些地區,通過持續的移除工作,已經成功控制了斑腿樹蛙的擴散。然而,外來入侵種的問題仍然嚴峻,需要長期的努力和多方面的合作才能有效解決。
台灣蛙類外來入侵種的問題不僅是一個生態問題,也是一個社會問題。它涉及到生態保護、經濟發展、公共教育等多個層面。只有通過政府、科研機構、保育組織和公眾的共同努力,才能有效應對這一挑戰,保護台灣珍貴的生物多樣性。
3.5 台灣熱門賞蛙地點
台灣以其豐富的生物多樣性聞名,而蛙類作為這個島嶼生態系統中的重要成員,吸引了眾多自然愛好者和生態攝影師的關注。近年來,賞蛙活動在台灣逐漸成為一種popular的生態旅遊項目,不僅為人們提供了親近自然的機會,也提高了公眾對蛙類保育的認識。
在台灣北部,陽明山國家公園是一個備受歡迎的賞蛙地點。這裡的多樣化地形和豐富的水資源為多種蛙類提供了理想的棲息環境。在陽明山,遊客可以在夜間聆聽到各種蛙鳴聲,其中最引人注目的是台北樹蛙的叫聲。這種台灣特有種的鳴叫聲類似「格格格」的重複音,在夜晚的山林中特別清晰。除了台北樹蛙,遊客還可能看到諸如面天樹蛙、莫氏樹蛙等物種。陽明山國家公園管理處還定期舉辦導覽活動,由專業解說員帶領遊客認識不同的蛙類及其生態。
在台北市區內,台北植物園也是一個意想不到的賞蛙勝地。儘管位於都市中心,這個擁有百年歷史的植物園仍然保留了豐富的水生植物和濕地環境,為多種蛙類提供了棲息地。在這裡,遊客可以近距離觀察到諸如澤蛙、小雨蛙等適應都市環境的蛙類。台北植物園的賞蛙活動特別適合家庭和兒童參與,因為這裡的環境相對安全,且交通便利。
向南移動到桃園,山地林班地(現已為拉拉山自然保護區的一部分)是觀察高海拔蛙類的絕佳地點。這裡的原始森林和清澈溪流為諸如梭德氏赤蛙、褐樹蛙等山區蛙類提供了理想的棲息環境。在夏季的夜晚,遊客可以聽到多種蛙類此起彼伏的鳴叫聲,構成了一場自然的交響樂。然而,由於地處偏遠且地形複雜,建議遊客參加專業導覽團,以確保安全並獲得更豐富的生態知識。
在台灣中部,日月潭國家風景區不僅以其優美的湖光山色聞名,也是一個excellent的賞蛙地點。環繞日月潭的森林和濕地為多種蛙類提供了棲息地。在這裡,遊客可能看到諸如腹斑蛙、白頷樹蛙等物種。特別是在春夏之交的繁殖季節,蛙類的活動更為頻繁,為觀察和攝影提供了絕佳機會。
台灣南部的墾丁國家公園是另一個熱門的賞蛙地點。這裡獨特的熱帶氣候孕育了豐富的蛙類資源,包括一些在台灣其他地方難得一見的物種,如史丹吉氏小雨蛙。墾丁的賞蛙活動通常集中在社頂自然公園和龍鑾潭自然中心等區域。這些地方不僅蛙類資源豐富,還有良好的步道系統和解說設施,方便遊客觀察和學習。
在台灣東部,花蓮的馬太鞍濕地是一個獨特的賞蛙地點。這片位於花東縱谷的濕地保存了豐富的水生生態系統,是多種蛙類的棲息地。在這裡,遊客可以觀察到諸如古氏赤蛙、澤蛙等喜好濕地環境的蛙類。馬太鞍濕地的賞蛙活動通常結合了阿美族文化體驗,讓遊客不僅能欣賞到大自然的奧妙,還能了解當地原住民與自然和諧共處的智慧。
值得一提的是,台灣的許多都市公園和校園也成為了便利的賞蛙地點。例如,台中都會公園、高雄都會公園等都市綠地,以及諸如東華大學、屏東科技大學等campus,都保留了相當豐富的蛙類資源。這些地方的賞蛙活動特別適合都市居民參與,讓人們不必遠行就能親近自然,認識蛙類。
然而,在推廣賞蛙活動的同時,也面臨著一些挑戰。首先是如何在滿足人們親近自然需求的同時,最小化對蛙類及其棲息地的干擾。許多賞蛙地點已經制定了strict的參觀規則,如限制使用強光手電筒、禁止捕捉和觸摸蛙類等。其次,如何提高賞蛙活動的教育價值也是一個重要issue。許多地方正在努力培訓專業的解說員,開發互動式的教育項目,以增進遊客對蛙類生態的了解。
另一個challenge是如何平衡保育需求和旅遊發展。一些popular的賞蛙地點面臨著遊客過多的問題,這可能對脆弱的生態系統造成壓力。為此,一些地方採取了限制遊客數量、實施預約制等措施,以控制遊客流量。
台灣的賞蛙活動不僅僅是一種休閒活動,更是重要的環境教育途徑。通過親身體驗,人們能夠更深入地了解蛙類的生態習性、面臨的威脅以及保育的重要性。許多參與賞蛙活動的人會產生對自然的敬畏之心,進而支持和參與到更廣泛的生態保育工作中。
此外,賞蛙活動也為科學研究提供了寶貴的資料。許多業餘自然愛好者通過長期觀察,為蛙類的分布、行為習性等研究貢獻了有價值的資訊。一些地方甚至開展了公民科學項目,鼓勵公眾參與到蛙類監測工作中來,這不僅擴大了監測的範圍和深度,也提高了公眾參與科學研究的興趣。

4. 蛙類解剖學與形態學
蛙類的解剖學與形態學是理解這些兩棲動物生理功能和適應性的基礎。蛙類的外部特徵包括光滑濕潤的皮膚、突出的眼睛和強壯的後腿,這些特徵都是為了適應其半水半陸的生活方式。蛙類的皮膚不僅是保護層,還是重要的呼吸器官,能夠透過皮膚進行氣體交換。
蛙類的骨骼系統經過特殊演化,以適應跳躍和游泳。他們的脊椎骨數量減少,骨盆與薦椎融合,形成堅固的薦骨,為強勁的後腿提供支撐。蛙類的頭骨結構也十分獨特,具有扁平的頭蓋骨和大型的眼窩,有利於捕捉獵物。
蛙類的感覺器官也有特殊適應。他們的眼睛能夠突出水面,同時觀察水上和水下環境。蛙類的中耳結構包括鼓膜和柱狀骨,能夠有效接收聲波,這對於他們的繁殖和通訊至關重要。一些蛙類還擁有特殊的觸覺器官,如手掌上的小疣,可以幫助他們感知周圍環境。
在解剖學上,蛙類還有一些獨特的適應性特徵。例如,某些樹蛙的腳趾末端有吸盤,能夠在光滑的樹葉和樹枝上攀爬。另外,一些蛙類的舌頭結構特殊,能夠快速彈出捕捉獵物,這種機制涉及複雜的肌肉和骨骼配合。
4.1 蛙類外部特徵
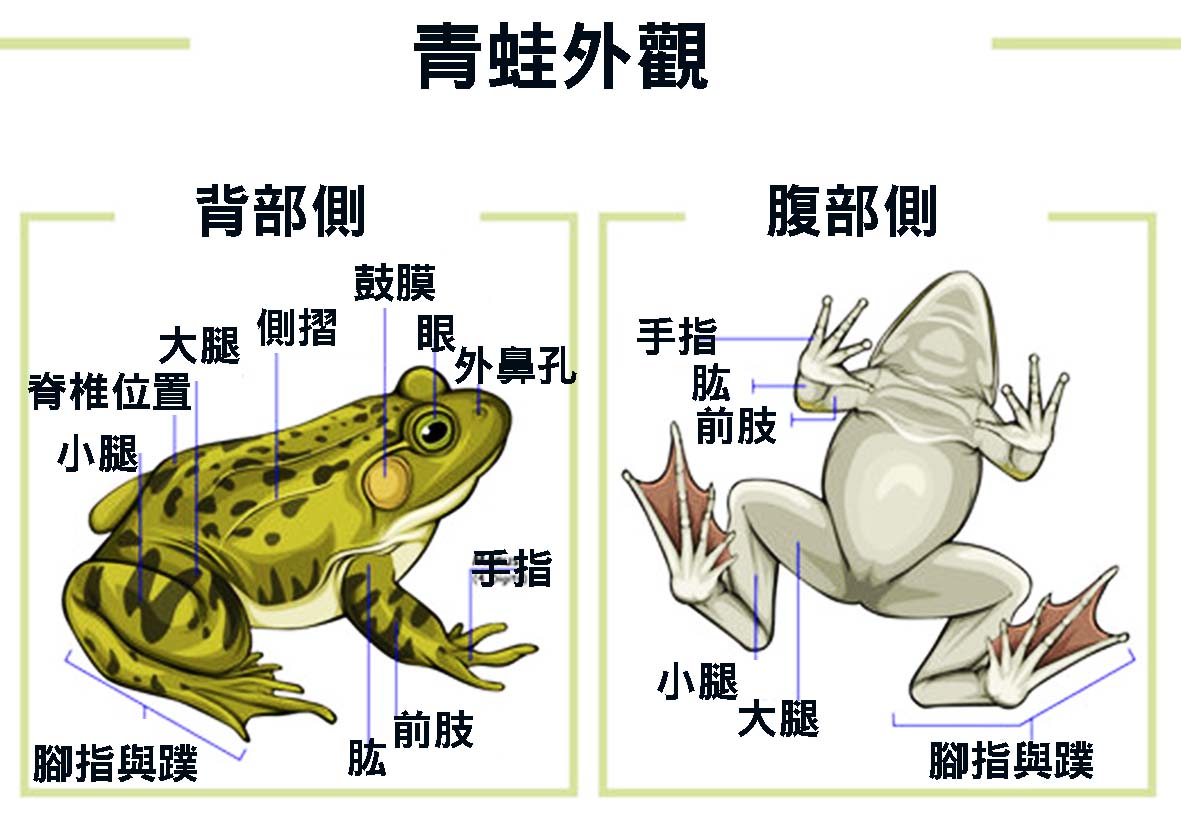
蛙類的外部特徵是其適應環境和生存的關鍵。這些特徵不僅反映了蛙類的進化歷程,也展現了其獨特的生理功能和生態適應性。蛙類的身體結構通常呈現扁平橢圓形,這種形態有助於減少水中阻力,同時也便於在陸地上快速移動。
蛙類最顯著的外部特徵之一是其光滑濕潤的皮膚。這層皮膚不僅具有保護功能,更是蛙類呼吸系統的重要組成部分。蛙類皮膚表面佈滿了微小的血管,能夠直接進行氣體交換,補充肺部呼吸。此外,皮膚還能分泌黏液,保持身體濕潤,這對於維持體內水分平衡至關重要。某些蛙類,如樹蛙,其皮膚還具有特殊的黏性,有助於它們在光滑的表面攀爬。
蛙類的頭部結構也十分獨特。他們擁有一個寬大而扁平的頭部,這種構造有利於增加口腔容積,提高捕食效率。蛙類的眼睛通常大而突出,位於頭部兩側,提供了廣闊的視野。這種眼睛結構不僅能夠在水面上方觀察周圍環境,還可以在水下保持警惕。許多蛙類的眼睛還具有第三眼瞼,稱為瞬膜(nictitating membrane),可以在水下保護眼睛並維持視力。
在頭部兩側,蛙類通常有明顯的圓形鼓膜。這是聽覺器官的外部結構,能夠接收聲波並傳導至內耳。鼓膜的大小和位置因種類而異,反映了不同蛙類對聲音感知的特化程度。某些樹蛙的鼓膜特別發達,這與其複雜的鳴叫行為有關。
蛙類的口部結構也值得關注。大多數蛙類擁有寬大的口裂,延伸至頭部兩側,有利於捕捉較大的獵物。口腔內部通常沒有牙齒,但上顎可能有一些小齒,主要用於固定獵物。蛙類的舌頭結構獨特,通常附著在口腔前部,可以快速彈出捕捉獵物。例如,綠樹蛙(Litoria caerulea)的舌頭長度可達其體長的三分之一,能在0.07秒內完成伸出和收回的動作。
蛙類的四肢結構展現了其適應多樣環境的能力。前肢相對較短,主要用於支撐身體和輔助移動。某些蛙類,如負子蛙(Ates obstetricans),前肢還具有特殊功能,用於攜帶卵串。後肢則明顯較長且強壯,適合跳躍和游泳。後腿的肌肉發達,能夠產生強大的推力,使蛙類能夠進行長距離跳躍。例如,牛蛙(Rana catesbeiana)能夠一躍跳出超過其體長6倍的距離。
蛙類的腳部結構也因應不同的生活環境而有所變化。水生性強的蛙類,如牛蛙,其腳趾間通常有蹼,增加了水中推進力。而樹棲蛙類,如紅眼樹蛙(Agachnis callidryas),其腳趾末端則有吸盤狀結構,能夠牢固地附著在樹葉或樹枝上。某些地棲蛙類,如負子蛙,腳掌上還有硬化的突起,有助於在陸地上行走和挖掘。
蛙類的皮膚顏色和圖案也是重要的外部特徵,往往與其生存環境密切相關。許多蛙類具有保護色,能夠與周圍環境融為一體,避免被捕食者發現。例如,綠色的樹蛙可以輕易地隱藏在樹葉間。而一些毒蛙,如箭毒蛙(Dendrobates)屬的物種,則擁有鮮艷的警戒色,警告潛在捕食者它們體內含有毒素。
某些蛙類還具有能夠改變體色的能力,這種特性稱為變色(metachrosis)。例如,樹蛙科的一些物種可以根據環境光線、溫度或情緒狀態改變體色。這種能力不僅有助於偽裝,還在體溫調節和社交行為中發揮作用。
此外,許多蛙類的皮膚上還有各種腺體,這些腺體可以分泌各種物質。最常見的是黏液腺,分泌保持皮膚濕潤的黏液。某些蛙類還擁有毒腺,分泌具有防禦功能的毒素。例如,蟾蜍(Bufo)屬的物種在頭部後方有明顯的腮腺,能夠分泌刺激性物質以驅趕捕食者。
在繁殖季節,許多雄性蛙類會發展出特殊的外部特徵。例如,某些蛙類的雄性會在前肢上長出婚姻墊(nuptial pad),這是一種角質化的結構,有助於在交配時緊抓雌性。另一些蛙類,如歐洲草蛙(Rana temporaria)的雄性,在繁殖季節會呈現出更鮮艷的體色。
值得注意的是,蛙類的外部特徵在其生命週期中會發生顯著變化。蝌蚪階段的蛙類外形與成體大不相同,通常呈現魚形,有尾巴和鰓,適應完全水生的生活方式。隨著變態過程的進行,蝌蚪逐漸發育出四肢,吸收尾巴,最終形成成體的形態。這種巨大的形態變化反映了蛙類從水生到半水半陸生活方式的適應過程。
4.2 蛙類骨骼系統
蛙類的骨骼系統是其身體結構的核心支撐,經過長期進化而形成獨特的適應性特徵。這個系統不僅為蛙類提供了身體的框架,還直接影響了它們的運動能力、生存策略和生態位。蛙類的骨骼結構反映了其半水半陸生活方式的需求,既能適應水中游泳,又能在陸地上靈活跳躍。
蛙類的頭骨結構是其骨骼系統中最為複雜的部分之一。頭骨扁平而寬大,這種構造有利於增加口腔容積,提高捕食效率。頭骨由多塊骨頭融合而成,包括額骨、頂骨、枕骨等。特別值得注意的是,蛙類的顎骨結構獨特,上顎骨和下顎骨能夠產生較大的開合動作,有助於捕捉較大的獵物。在頭骨兩側,還有明顯的眼眶,為突出的大眼睛提供了保護和支撐。
蛙類的脊椎骨數量相對較少,通常只有9節,這是對跳躍生活方式的適應。相比之下,蠑螈等其他兩棲動物的脊椎骨數量要多得多。蛙類的脊椎骨中,第一節稱為環椎,與頭骨相連;最後一節則與尾椎骨融合,形成尾骨。值得注意的是,蛙類的薦椎(通常是第9節脊椎)特別發達,兩側延伸出翼狀突起,與髂骨相連,形成了堅固的薦骨。這種結構為強壯的後肢提供了穩固的支撐點,是蛙類能夠進行有力跳躍的關鍵。
蛙類的肩帶結構也很特殊。它由肩胛骨、鎖骨和匙骨組成,形成了一個堅固的環狀結構,為前肢的活動提供了支撐。這種結構允許前肢在多個方向上自由活動,有利於爬行、游泳和捕食。值得一提的是,蛙類的胸骨結構相對簡單,這與其獨特的呼吸方式有關。
蛙類的前肢骨骼結構相對簡單,包括肱骨、橈骨和尺骨。這些骨骼較短,主要用於支撐身體和輔助移動。前肢的掌骨和指骨結構因應不同種類的生活環境而有所變化。例如,樹蛙的指骨末端通常會擴張,形成吸盤狀結構,有助於在樹枝或樹葉上攀爬。
相比之下,蛙類的後肢骨骼結構更為發達和複雜,這是其跳躍能力的基礎。後肢由股骨、脛腓骨(由脛骨和腓骨融合而成)以及跗骨和趾骨組成。股骨與脛腓骨之間的關節允許大幅度的屈伸運動,這是蛙類能夠進行長距離跳躍的關鍵。跗骨特別延長,形成了額外的槓桿,進一步增強了跳躍的力量。後肢的趾骨通常比前肢的長,有些水生蛙類的趾間還有蹼,增加了水中的推進力。
蛙類的骨盆結構也很獨特。髂骨、坐骨和恥骨融合形成了堅固的骨盆腔,為強壯的後肢肌肉提供了附著點。髂骨特別延長,與薦椎相連,形成了一個V字形的結構,這種構造能夠有效地將後肢的力量傳遞到整個身體。
值得注意的是,蛙類的骨骼系統在其生命週期中經歷了巨大的變化。蝌蚪階段的骨骼結構與成體完全不同,主要由軟骨組成,適應完全水生的生活方式。隨著變態過程的進行,軟骨逐漸被骨組織替代,同時發育出四肢骨骼,尾部骨骼則逐漸退化。這種戲劇性的變化反映了蛙類從水生到半水半陸生活方式的適應過程。
蛙類骨骼系統的另一個重要特徵是其獨特的骨質結構。與哺乳動物不同,蛙類的骨骼通常是中空的,這種結構在保證強度的同時大大減輕了體重,有利於跳躍和游泳。此外,蛙類的骨骼中含有豐富的鈣質存儲區,這在它們的新陳代謝和鈣平衡中起著重要作用。
某些蛙類還發展出了特殊的骨骼適應。例如,某些樹蛙的指骨和趾骨間關節特別靈活,允許它們的腳掌能夠在多個方向彎曲,這有助於它們在複雜的樹冠環境中移動。另一個例子是某些地棲蛙類,如負子蛙,它們的骨盆結構特別寬大,為攜帶卵串提供了空間。
蛙類的頭骨結構中還包括了聽覺系統的重要部分。在內耳區域,有一種特殊的骨頭稱為柱狀骨(columella),它連接鼓膜和內耳,負責將聲波從外界傳導到內耳。這種結構使得蛙類能夠敏銳地感知環境中的聲音,對於它們的交流和捕食至關重要。
在某些蛙類中,骨骼系統還與防禦機制密切相關。例如,某些樹蛙的頭骨和脊椎骨上有鋒利的骨質突起,這些突起穿透皮膚,形成了有效的防禦武器。另一個有趣的例子是非洲毛蛙(Trichobatrachus robustus),雄性個體在繁殖季節會發展出類似爪子的骨質結構,這些「爪子」實際上是斷裂的趾骨,穿透皮膚後形成了防禦和競爭的武器。
蛙類的骨骼系統還與它們的呼吸機制密切相關。由於蛙類缺乏肋骨,它們主要依靠口腔底部的肌肉運動來進行肺部呼吸。這種特殊的呼吸方式被稱為鼓式呼吸(buccal pumping),它不僅影響了蛙類的骨骼結構,也塑造了它們獨特的生理特徵。
4.3 蛙類皮膚結構與功能
蛙類的皮膚是一個極其複雜且多功能的器官,在蛙類的生理和生態適應中扮演著至關重要的角色。這層看似簡單的外層不僅是蛙類身體的第一道防線,還承擔著呼吸、水分調節、體溫控制等多項重要功能。蛙類皮膚的獨特結構和功能是它們能夠在水陸兩棲環境中成功生存的關鍵因素之一。
蛙類皮膚的基本結構由表皮和真皮兩層組成。表皮是最外層,主要由角質化的細胞構成,這些細胞定期脫落和更新,保持皮膚的健康狀態。真皮層位於表皮之下,含有豐富的血管、神經末梢和各種腺體。這兩層結構緊密結合,共同發揮著保護和生理調節的功能。
蛙類皮膚最顯著的特徵之一是其高度的通透性。與其他陸生脊椎動物不同,蛙類的皮膚缺乏角質層,這使得它們的皮膚更容易滲透水分和氣體。這種結構雖然增加了脫水的風險,但也為蛙類提供了獨特的呼吸能力。蛙類的皮膚表面佈滿了微小的血管,能夠直接進行氣體交換,這種被稱為皮膚呼吸的過程對蛙類的生存至關重要。某些完全水生的蛙類,如非洲爪蟾(Xenopus laevis),甚至可以完全依賴皮膚呼吸生存。
蛙類皮膚的另一個重要功能是維持體內水分平衡。由於皮膚的高通透性,蛙類面臨著持續的水分流失風險。為了應對這一挑戰,蛙類皮膚上分佈著大量的黏液腺。這些腺體分泌黏液,在皮膚表面形成一層保護膜,減少水分蒸發。同時,蛙類還能夠通過皮膚主動吸收環境中的水分。例如,當處於乾燥環境時,蛙類會採取特殊的姿勢,將腹部緊貼潮濕的地面,通過皮膚吸收水分。這種能力使得蛙類能夠在相對乾燥的環境中生存。
蛙類的皮膚還在體溫調節中發揮著關鍵作用。作為變溫動物,蛙類主要依靠環境來調節體溫。它們的皮膚富含血管,可以通過調節皮膚血流量來控制熱量散失或吸收。當環境溫度過高時,蛙類會增加皮膚血流量,促進熱量散失;反之,當環境溫度較低時,蛙類會減少皮膚血流量,以保存體熱。某些樹蛙,如澳大利亞綠樹蛙(Litoria caerulea),甚至能夠通過改變皮膚顏色來調節吸收的熱量。
蛙類皮膚的另一個重要功能是分泌各種化學物質。除了保持皮膚濕潤的黏液外,許多蛙類還能分泌具有抗菌、抗真菌或毒性的物質。這些分泌物在蛙類的防禦機制中扮演著重要角色。例如,某些樹蛙分泌的蠟狀物質可以防止水分流失,而一些地棲蛙類則分泌具有強烈刺激性的物質來驅趕捕食者。最著名的例子莫過於箭毒蛙(Dendrobates),它們的皮膚能夠分泌強烈的神經毒素,這些毒素不僅能夠有效地抵禦捕食者,還被某些南美原住民用來淬毒箭矢。
蛙類皮膚的顏色和圖案也是其重要特徵,這些特徵往往與環境適應和行為密切相關。許多蛙類擁有保護色,能夠與周圍環境融為一體。例如,生活在樹林中的綠樹蛙通常呈現綠色,而生活在地面的蛙類則常常呈現褐色或灰色。一些蛙類還具有變色能力,可以根據環境、情緒狀態或繁殖季節改變體色。這種能力不僅有助於偽裝,還在社交行為和溫度調節中發揮作用。
蛙類皮膚還具有感知功能。皮膚表面分佈著大量的感受器,能夠感知溫度、壓力和化學刺激。這些感受器使蛙類能夠迅速感知環境變化和潛在威脅。某些蛙類,如樹蛙,腳掌上還有特殊的觸覺感受器,有助於它們在樹枝或樹葉上安全地移動和攀爬。
在某些蛙類中,皮膚還承擔著繁殖相關的功能。例如,智利袋蟾(Rhinoderma darwinii)的雄性會將受精卵儲存在聲囊中孵化。這種特殊的育幼方式需要聲囊內部的皮膚具有特殊的結構,能夠為發育中的幼體提供氧氣和營養。
蛙類皮膚的另一個有趣特徵是其再生能力。與哺乳動物不同,蛙類能夠快速且完全地修復皮膚損傷,而不留疤痕。這種能力不僅有助於恢復受損的保護屏障,還能維持皮膚的生理功能。科學家們正在研究蛙類皮膚的再生機制,希望能為人類醫學帶來啟發。
值得注意的是,蛙類的皮膚結構和功能在其生命週期中會發生顯著變化。蝌蚪階段的皮膚結構相對簡單,主要適應水生環境。隨著變態過程的進行,皮膚結構逐漸複雜化,發展出適應陸地生活的特徵。這種轉變包括皮膚腺體的發育、色素細胞的分化以及感受器的形成等。
蛙類皮膚的這些獨特特性使其成為生物學研究的重要對象。科學家們正在深入研究蛙類皮膚的分子機制,希望能夠開發新的藥物或材料。例如,某些蛙類皮膚分泌物中發現的抗菌肽已經成為新型抗生素研究的重要方向。同時,蛙類皮膚對環境變化的敏感性也使其成為環境監測的重要指標,在生態保護中發揮著重要作用。
4.4 蛙類感覺器官
蛙類的感覺器官是其適應環境和生存的關鍵工具,它們經過長期進化,形成了一套高度專門化的感知系統。這些感覺器官不僅幫助蛙類捕捉獵物、躲避天敵,還在其社交行為和繁殖活動中扮演著重要角色。蛙類的主要感覺器官包括視覺、聽覺、嗅覺、味覺和觸覺系統,每一種感覺器官都有其獨特的結構和功能。
視覺是蛙類最為發達的感覺之一。蛙類的眼睛通常大而突出,位於頭部兩側,提供了近乎360度的視野。這種廣闊的視野對於捕捉快速移動的獵物和察覺潛在威脅至關重要。蛙類的眼睛結構與其他脊椎動物相似,包括角膜、虹膜、晶狀體和視網膜。然而,蛙類眼睛有幾個獨特之處。首先,蛙類的晶狀體呈扁平狀,這種結構使得蛙類在水下和空氣中都能保持良好的視力。其次,蛙類擁有一種特殊的眼瞼,稱為瞬膜(nictitating membrane),這是一層透明的膜,可以在蛙類潛水時保護眼睛,同時允許視力不受影響。
蛙類的視覺系統還具有出色的顏色感知能力。許多蛙類能夠識別各種顏色,這對於辨認同類、尋找食物和識別潛在威脅都非常重要。例如,某些樹蛙能夠通過顏色識別來選擇最適合的棲息地。值得注意的是,一些夜行性蛙類的視網膜結構有所不同,具有更多的桿狀細胞,這使它們能在低光環境下保持良好的視力。
聽覺是蛙類另一個高度發達的感官系統。蛙類的耳朵結構獨特,沒有外耳,而是在頭部兩側有一個圓形的鼓膜。聲波通過鼓膜傳導到中耳的柱狀骨(columella),然後傳遞到內耳。蛙類的聽覺系統對特定頻率的聲音特別敏感,這些頻率通常與同類的叫聲相對應。這種特化使得蛙類能夠在嘈雜的環境中準確識別同類的叫聲,這在繁殖季節尤為重要。
某些蛙類,如樹蛙,還發展出了特殊的聽覺適應。它們的中耳腔與口腔相連,形成了一個聲音共振腔,這種結構增強了某些頻率的聲音,使得樹蛙能夠更好地在複雜的森林環境中進行聲音通訊。此外,一些蛙類還能夠通過身體其他部位感知聲波振動,這種能力補充了傳統的聽覺系統。
嗅覺在蛙類的生活中也扮演著重要角色,儘管相比視覺和聽覺,蛙類的嗅覺系統相對簡單。蛙類的鼻腔內有嗅上皮,能夠感知空氣中的化學物質。有趣的是,蛙類有兩套嗅覺系統:主嗅覺系統和副嗅覺系統(也稱為犁鼻器)。主嗅覺系統主要用於感知一般的氣味,而副嗅覺系統則專門用於感知費洛蒙等特殊化學信號。這種雙重嗅覺系統在蛙類的繁殖行為中尤為重要,幫助它們識別和吸引配偶。
味覺是蛙類感官系統中相對不太發達的一部分。蛙類的舌頭上有味蕾,但數量遠少於哺乳動物。然而,這並不意味著味覺對蛙類不重要。事實上,蛙類的味覺系統與其捕食行為密切相關。當蛙類捕獲獵物時,它們會通過舌頭上的味蕾快速判斷食物的適口性。一些研究表明,某些蛙類能夠通過味覺辨別有毒和無毒的獵物,這對於避免攝入有害物質至關重要。
觸覺是蛙類另一個重要的感官系統。蛙類的皮膚上分佈著大量的觸覺感受器,能夠感知壓力、振動和溫度變化。這些感受器在蛙類的日常活動中發揮著重要作用,幫助它們感知周圍環境的變化。例如,當捕食者接近時,蛙類能夠通過皮膚感受到水或地面的微小振動,從而迅速做出反應。
某些蛙類還發展出了特殊的觸覺適應。例如,許多樹蛙的腳掌上有特殊的觸覺感受器,這些感受器能夠幫助它們在光滑的樹葉或樹枝上安全地移動和攀爬。另一個有趣的例子是非洲爪蟾(Xenopus laevis),它們的前肢上有特殊的觸鬚,這些觸鬚能夠在濁水中感知周圍環境,幫助它們尋找食物。
值得注意的是,蛙類的感覺器官在其生命週期中會經歷顯著變化。蝌蚪階段的感官系統相對簡單,主要適應水生環境。例如,蝌蚪階段的視覺系統主要用於感知光線強度和方向,而不是形狀和顏色。隨著變態過程的進行,蛙類的感官系統逐漸發育成熟,適應陸地生活的需求。
蛙類的感覺器官還展現出驚人的適應性。例如,某些生活在洞穴環境中的蛙類,如墨西哥洞穴蛙(Eleutherodactylus cisternasii),其視覺系統退化,但聽覺和觸覺系統變得更加敏銳。這種感官系統的重組反映了蛙類對特定環境的適應能力。
此外,一些蛙類還具有特殊的感知能力。例如,某些樹蛙能夠感知紫外線,這種能力可能有助於它們在複雜的森林環境中辨識同類或獵物。另一個例子是某些水生蛙類,如非洲爪蟾,它們具有側線系統,這是一種通常見於魚類的感覺器官,能夠感知水流的變化和水中物體的移動。
蛙類的感覺器官不僅在個體生存中發揮重要作用,還在種群和生態系統層面具有重要意義。例如,蛙類的聽覺系統使它們能夠通過鳴叫進行種內交流,這種交流對於維持種群結構和繁衍後代至關重要。同時,蛙類敏感的感官系統也使它們成為環境變化的重要指示器,在生態監測和保護中發揮著重要作用。
4.5 蛙類解剖學的獨特適應
蛙類在長期的進化過程中形成了許多獨特的解剖學適應,這些適應使得蛙類能夠成功地生存在多樣化的環境中,從熱帶雨林到乾旱沙漠,從高山湖泊到城市公園。這些解剖學特徵不僅反映了蛙類的進化歷史,也展現了它們對不同生態位的精妙適應。
蛙類最顯著的解剖學適應之一是其獨特的口腔和舌頭結構。與大多數脊椎動物不同,蛙類的舌頭前端附著在口腔前部,而後端則是自由的。這種結構使得蛙類能夠快速彈出舌頭捕捉獵物,整個過程可能只需要0.07秒。舌頭表面覆蓋著黏液,有助於粘住獵物。更令人驚訝的是,某些蛙類的舌頭長度可達其體長的1.4倍。例如,角舌蛙(Hemiphractus proboscideus)擁有相對體型最長的舌頭,這種適應使它們能夠捕捉遠處的獵物,大大擴展了其覓食範圍。
蛙類的眼睛結構也具有獨特的適應性。大多數蛙類擁有突出的大眼睛,這不僅提供了寬廣的視野,還能夠在進食時幫助將食物推入喉嚨。蛙類的眼球能夠收縮到頭骨內,這種機制在吞嚥食物時特別有用,因為突出的眼球可以向下壓迫口腔,協助食物的運輸。此外,許多蛙類還擁有一種名為瞬膜的特殊眼瞼,這層透明的膜能夠在蛙類潛水或跳躍時保護眼睛,同時不影響視力。
蛙類的耳朵結構也是其解剖學適應的一個典型例子。蛙類沒有外耳,而是在頭部兩側有一個圓形的鼓膜。這種結構不僅能夠有效接收聲波,還能減少水中運動時的阻力。更有趣的是,某些樹蛙的中耳腔與口腔相連,形成了一個聲音共振腔。這種適應增強了特定頻率的聲音,使得這些蛙類能夠在嘈雜的環境中更好地進行聲音通訊。
蛙類的骨骼系統也展現了許多獨特的適應。最顯著的是其拉長的後肢骨骼,這是蛙類強大跳躍能力的基礎。蛙類的跳躍不僅依賴於強壯的肌肉,還得益於其特殊的骨骼結構。例如,蛙類的跗骨特別延長,形成了額外的槓桿,進一步增強了跳躍的力量。同時,蛙類的脊椎骨數量相對較少,通常只有9節,這種結構增加了身體的靈活性,有利於跳躍和游泳。
蛙類的皮膚結構是另一個重要的解剖學適應。與其他陸生脊椎動物不同,蛙類的皮膚缺乏角質層,這使得它們的皮膚具有高度的通透性。這種特性雖然增加了脫水的風險,但也為蛙類提供了獨特的呼吸能力。蛙類的皮膚表面佈滿了微小的血管,能夠直接進行氣體交換,這種被稱為皮膚呼吸的過程在某些完全水生的蛙類中尤為重要。例如,非洲爪蟾(Xenopus laevis)能夠完全依賴皮膚呼吸生存。
某些蛙類還發展出了特殊的皮膚腺體,這些腺體能夠分泌各種化學物質,用於防禦、交流或維持體內平衡。例如,某些樹蛙分泌的蠟狀物質可以防止水分流失,而一些地棲蛙類則分泌具有強烈刺激性的物質來驅趕捕食者。最極端的例子莫過於箭毒蛙(Dendrobates),它們的皮膚能夠分泌強烈的神經毒素,這些毒素不僅能夠有效地抵禦捕食者,還被某些南美原住民用來淬毒箭矢。
蛙類的循環系統也表現出獨特的適應。與其他脊椎動物不同,成年蛙類擁有三室心臟(兩心房一心室),而不是四室心臟。這種結構雖然允許一定程度的血液混合,但也為蛙類提供了更大的生理靈活性,使它們能夠適應不同的環境條件。例如,當蛙類潛水時,它們可以將血液流向皮膚增加,以提高皮膚呼吸的效率。
蛙類的消化系統也有一些獨特的適應。許多蛙類擁有可伸縮的胃,這使得它們能夠一次性吞食相對較大的獵物。此外,蛙類的肝臟能夠產生尿素,這種物質可以幫助蛙類在乾旱環境中保持體內水分平衡。例如,某些生活在乾旱地區的蛙類,如澳大利亞的水囊蛙(Cyclorana platycephala),能夠在乾季時將尿素儲存在體內,以維持必要的水分。
某些蛙類還發展出了特殊的繁殖相關的解剖學適應。例如,智利袋蟾(Rhinoderma darwinii)的雄性擁有特化的聲囊,用於孵化卵和幼體。這種特殊的育幼方式需要聲囊內部的皮膚具有特殊的結構,能夠為發育中的幼體提供氧氣和營養。另一個例子是蘇利南海蟾(Pipa pipa),雌性的背部皮膚會形成小窩,用於孵化卵和幼體。
蛙類的呼吸系統也展現了獨特的適應。除了上述提到的皮膚呼吸外,蛙類還發展出了一種特殊的肺部呼吸機制,稱為鼓式呼吸(buccal pumping)。由於蛙類缺乏肋骨,它們主要依靠口腔底部的肌肉運動來進行肺部呼吸。這種呼吸方式不僅適應了蛙類的身體結構,還使得蛙類能夠在水下長時間屏住呼吸。
某些蛙類還發展出了特殊的運動相關的解剖學適應。例如,某些樹蛙的腳趾末端有特殊的吸盤結構,這些吸盤不僅有助於攀爬,還能分泌特殊的黏液,增強附著力。另一個有趣的例子是飛蛙(Rhacophorus),它們的腳趾間有特大的蹼,這種結構使得它們能夠在空中滑翔,有效地擴大了活動範圍。
這些獨特的解剖學適應不僅使蛙類能夠在各種環境中生存,還使它們成為生物學研究的重要對象。通過研究蛙類的這些適應性特徵,科學家們不僅能夠更好地理解進化過程,還可能為生物醫學、材料科學等領域帶來新的啟發。

5. 蛙類生理學與適應
蛙類的生理學和適應機制是它們能夠在多樣化環境中生存的關鍵。作為兩棲動物,蛙類面臨著水陸兩棲生活的挑戰,因此發展出了一系列獨特的生理適應。
蛙類的滲透調節能力尤為突出。它們能夠通過皮膚和膀胱調節體內水分和離子平衡,這使得蛙類能夠在淡水和陸地環境中維持體內穩態。例如,某些樹蛙能夠在乾燥環境中通過皮膚分泌特殊的蠟質物質減少水分流失。
體溫調節是蛙類另一個重要的生理適應。作為變溫動物,蛙類主要依賴行為和環境來調節體溫。它們可以通過改變體位、選擇適宜的微環境來維持最佳體溫。某些蛙類甚至能夠通過改變皮膚顏色來調節熱量吸收。
蛙類的變態過程是其生理適應的一個顯著特徵。從水生的蝌蚪到陸生的成蛙,這一過程涉及複雜的形態和生理變化,包括呼吸系統、循環系統和消化系統的重組。這種生理上的巨大轉變使得蛙類能夠適應不同的生態位。
蛙類的呼吸機制也展現了獨特的適應性。除了肺部呼吸,許多蛙類還能通過皮膚進行氣體交換,這種能力在某些完全水生的蛙類中尤為重要。此外,蛙類的鼓式呼吸(buccal pumping)機制也是其適應陸地生活的重要特徵。
5.1 蛙類滲透調節
蛙類的滲透調節是其生理適應中最為關鍵的機制之一,它使得這些兩棲動物能夠在水陸環境之間自如轉換。滲透調節是指生物體維持體內水分和電解質平衡的過程,對於蛙類這樣的兩棲動物來說,這一機制尤為重要,因為它們經常面臨淡水環境中過度吸水和陸地環境中脫水的雙重挑戰。
在淡水環境中,蛙類面臨的主要問題是過度吸水。由於滲透作用,水分會不斷從環境中進入蛙體內。為了應對這一挑戰,蛙類演化出了一套高效的排水機制。首先,蛙類的皮膚具有選擇性滲透的特性,能夠在一定程度上控制水分的進入。其次,蛙類的腎臟能夠產生大量稀釋的尿液,迅速排出多餘的水分。例如,綠蛙(Rana clamitans)在淡水中能夠每天產生相當於其體重30-50%的尿液,這種高效的排水能力使其能夠維持體內的離子平衡。
除了腎臟,蛙類還擁有一個獨特的器官 - 膀胱,在滲透調節中發揮重要作用。蛙類的膀胱不僅是儲存尿液的場所,還能夠重新吸收水分和某些離子。當蛙類離開水環境進入陸地時,膀胱中儲存的水分成為其維持體內水平衡的重要來源。例如,某些沙漠蛙類,如澳大利亞的水囊蛙(Cyclorana platycephala),能夠在膀胱中儲存相當於其體重一半的水分,這使得它們能夠在乾旱環境中存活數月之久。
在陸地環境中,蛙類面臨的主要挑戰是防止脫水。為此,蛙類發展出了多種策略。首先,蛙類可以通過改變皮膚的通透性來減少水分流失。某些樹蛙,如澳大利亞綠樹蛙(Litoria caerulea),能夠分泌一種特殊的蠟質物質覆蓋在皮膚表面,大大減少了水分蒸發。其次,蛙類能夠通過行為調節來減少水分流失,如選擇濕度較高的微環境或者採取特殊的姿勢減少暴露在空氣中的體表面積。
蛙類的滲透調節還涉及到離子平衡的維持。在淡水環境中,蛙類面臨著體內鹽分流失的問題。為了補充失去的離子,蛙類演化出了一種特殊的器官 - 鹽泵(salt pump)。這種器官位於蛙類的皮膚表面,能夠主動從環境中吸收鈉離子和氯離子。例如,非洲爪蟾(Xenopus laevis)的皮膚上就分佈著大量的鹽泵,使其能夠在離子濃度極低的環境中維持體內的電解質平衡。
蛙類的滲透調節能力在其生命週期的不同階段也表現出顯著差異。蝌蚪階段的蛙類主要生活在水中,其滲透調節機制與成蛙有所不同。蝌蚪的皮膚通透性較高,主要依靠鰓和皮膚進行離子交換。隨著變態過程的進行,蝌蚪的滲透調節機制逐漸轉變為成蛙的模式,這個過程涉及到腎臟功能的發育、皮膚結構的改變以及新的內分泌調節機制的建立。
值得注意的是,蛙類的滲透調節還受到內分泌系統的調控。抗利尿激素(ADH)和醛固酮(aldosterone)是參與蛙類滲透調節的兩種重要激素。抗利尿激素可以增加腎小管和膀胱對水的重吸收,而醛固酮則促進鈉離子的重吸收。這些激素的分泌受到體內水分和電解質狀態的調控,形成了一個精密的反饋系統。
某些蛙類還發展出了特殊的滲透調節策略來應對極端環境。例如,某些生活在高鹽環境中的蛙類,如蟾蜍科的鹽池蟾蜍(Bufo viridis),能夠忍受體液滲透壓的顯著升高。這種適應使得它們能夠在鹽度較高的水體中生存。另一個極端的例子是某些生活在寒冷地區的蛙類,如林蛙(Rana sylvatica),它們能夠在體內積累大量的葡萄糖,作為一種天然的「防凍劑」,使得它們能夠在體液部分結冰的情況下存活下來。
蛙類的滲透調節能力還與其棲息地選擇密切相關。不同種類的蛙類根據其滲透調節能力選擇了不同的生態位。例如,擁有較強滲透調節能力的蛙類往往能夠在更為乾旱或者鹽度較高的環境中生存,而滲透調節能力較弱的種類則可能限制在濕度較高的環境中。這種生理特性與生態分佈的關聯為我們理解蛙類的地理分佈和進化歷史提供了重要線索。
蛙類的滲透調節機制不僅是其生存的關鍵,也為人類在醫學和工程領域帶來了啟發。例如,研究蛙類皮膚的選擇性滲透特性,可能為開發新型的人工膜或者藥物傳遞系統提供思路。而蛙類在極端環境下的滲透調節策略,也為我們探索生物在惡劣條件下的生存機制提供了寶貴的案例。
5.2 蛙類體溫調節
蛙類作為變溫動物,其體溫調節機制與恆溫動物有顯著不同。蛙類的體溫主要受環境溫度影響,但這並不意味著它們完全受制於環境。事實上,蛙類已經進化出一系列複雜的生理和行為策略來調節體溫,以適應多變的環境條件。
蛙類的體溫調節首先依賴於行為調節。這包括選擇合適的微環境、改變體位以及調整活動時間等。許多蛙類會根據環境溫度的變化在不同的微環境之間移動。例如,在寒冷的早晨,蛙類可能會選擇陽光直射的地方來提高體溫;而在炎熱的中午,它們則可能躲在陰涼的樹葉下或潛入水中以避免過熱。
蛙類還能通過改變體位來調節熱量的吸收和散失。當需要提高體溫時,蛙類可能會採取扁平的姿勢,增加與環境接觸的表面積,以加速熱量吸收。相反,當需要降低體溫時,它們可能會豎起身體,減少與熱源的接觸。例如,澳大利亞的綠樹蛙(Litoria caerulea)在寒冷時會展開四肢,將身體緊貼在溫暖的表面上,以最大限度地吸收熱量。
某些蛙類還能通過改變皮膚顏色來調節體溫。皮膚顏色的變化不僅用於偽裝和社交,還能影響熱量的吸收和反射。例如,某些樹蛙在寒冷時會變成較深的顏色以增加熱量吸收,而在高溫時則變為較淺的顏色以反射更多熱量。這種能力被稱為生理性變色(physiological color change),是蛙類體溫調節的一個重要方面。
蛙類的體溫調節還涉及到生理機制的調整。雖然蛙類不能像哺乳動物那樣產生大量的體內熱,但它們可以通過調整新陳代謝率來影響體溫。在寒冷環境中,某些蛙類能夠短暫地提高代謝率,產生一些額外的熱量。這種機制雖然不能長期維持,但可以幫助蛙類度過短暫的低溫期。
蛙類的皮膚在體溫調節中也扮演著重要角色。蛙類的皮膚富含血管,可以通過調節皮膚血流量來控制熱量散失或吸收。當環境溫度較高時,蛙類會增加皮膚血流量,促進熱量散失;當環境溫度較低時,則會減少皮膚血流量以保存體熱。此外,蛙類還可以通過蒸發冷卻來降低體溫。它們能夠分泌皮膚黏液,通過蒸發來帶走熱量,這種機制在高溫環境中特別重要。
某些蛙類還發展出了特殊的體溫調節策略來應對極端環境。例如,生活在寒冷地區的林蛙(Rana sylvatica)能夠在體內積累大量的葡萄糖,作為一種天然的「防凍劑」。這使得它們能夠在體液部分結冰的情況下存活下來,體溫可以降到零度以下而不致死亡。這種極端的適應能力使得林蛙能夠在北美寒冷的地區生存。
另一個極端的例子是生活在沙漠環境中的蛙類。澳大利亞的水囊蛙(Cyclorana platycephala)能夠在乾旱季節鑽入地下,分泌一層黏液包裹全身,形成一個保護性的茧。在這個茧中,水囊蛙可以降低代謝率,進入一種類似休眠的狀態,這種狀態可以持續數月之久,直到雨季來臨。這種策略不僅幫助水囊蛙度過高溫乾旱的環境,還能有效調節體溫,避免過熱和脫水。
蛙類的體溫調節能力在其生命週期的不同階段也表現出顯著差異。蝌蚪階段的蛙類主要生活在水中,其體溫調節主要依賴於水溫的變化。隨著變態過程的進行,蛙類逐漸獲得了更複雜的體溫調節能力,包括行為調節和生理調節。這個過程涉及到皮膚結構的改變、新的內分泌調節機制的建立,以及行為模式的轉變。
值得注意的是,蛙類的體溫調節還與其他生理過程密切相關。例如,蛙類的免疫系統功能與體溫有關,某些病原體在特定溫度範圍內更易繁殖。因此,蛙類通過調節體溫來增強免疫防禦。這種現象被稱為「行為性發熱」(behavioral fever),已在多種蛙類中被觀察到。
蛙類的體溫調節能力還與其生態位密切相關。不同種類的蛙類根據其體溫調節能力選擇了不同的棲息地和活動模式。例如,具有較強體溫調節能力的蛙類可能更容易適應溫度變化大的環境,而體溫調節能力較弱的種類則可能限制在溫度相對穩定的環境中。這種生理特性與生態分佈的關聯為我們理解蛙類的地理分佈和進化歷史提供了重要線索。
蛙類的體溫調節機制不僅是其生存的關鍵,也為人類在生物醫學和工程領域帶來了啟發。例如,研究蛙類如何在低溫環境中保護細胞免受凍傷,可能為開發新的器官保存技術提供思路。而蛙類在高溫環境下的降溫策略,也可能為設計新型的散熱系統提供靈感。
人類活動導致的氣候變化對蛙類的體溫調節提出了新的挑戰。全球變暖可能會改變蛙類棲息地的溫度模式,這可能超出某些蛙類的體溫調節能力範圍。因此,了解蛙類的體溫調節機制不僅有助於我們理解這些生物的生理特性,還對評估和預測氣候變化對蛙類種群的影響至關重要。
5.3 蛙類的變態
蛙類的變態是一個複雜而引人入勝的生物學過程,它標誌著蛙類從完全水生的蝌蚪階段轉變為能夠在陸地生活的成體。這個過程不僅涉及外部形態的巨大變化,還包括內部器官系統的重組和生理功能的轉變。變態過程是蛙類生命週期中最為關鍵的階段之一,反映了蛙類適應不同生態環境的演化策略。
變態過程通常始於蝌蚪後期,當蝌蚪達到一定大小和發育階段時,體內的內分泌系統開始產生變化。這個過程主要受甲狀腺激素的調控,特別是三碘甲狀腺原氨酸(T3)的分泌增加觸發了一系列的形態和生理變化。除了甲狀腺激素,其他激素如皮質醇和生長激素也在變態過程中發揮重要作用。
變態的第一個可見跡象通常是後肢的出現。後肢最初以小芽的形式在蝌蚪尾部附近出現,然後逐漸發育成完整的腿。有趣的是,後肢的發育過程是漸進的,而前肢則在變態後期突然出現。這是因為前肢在早期就已經在蝌蚪的鰓蓋下發育,只是在變態後期才突破鰓蓋露出。
隨著四肢的發育,蝌蚪的尾巴開始萎縮。尾巴的萎縮過程涉及細胞程序性死亡(細胞凋亡)和組織重吸收。有趣的是,尾巴中的物質並非完全被浪費,而是被重新利用於其他器官的發育。例如,尾巴中的肌肉蛋白可能被用於發育中的四肢肌肉。
在外部形態變化的同時,蛙類的內部器官系統也在經歷巨大的重組。呼吸系統從鰓呼吸轉變為肺呼吸是最顯著的變化之一。蝌蚪的鰓逐漸退化,同時肺部開始發育。有趣的是,在這個過程中,許多蛙類還發展出了皮膚呼吸的能力,這在某些水生性強的蛙類中尤為重要。
消化系統也經歷了重大的改變。蝌蚪主要以植物性食物為主,擁有相對較長的腸道。隨著變態的進行,腸道逐漸縮短,適應成蛙以昆蟲等動物性食物為主的飲食習慣。肝臟和胰臟等消化器官也隨之改變功能,以適應新的代謝需求。
循環系統同樣經歷了重組。蝌蚪階段的心臟是二腔心臟,隨著變態過程,逐漸發展為成蛙的三腔心臟。血液循環路徑也隨之改變,以適應從水生到陸生的生活方式轉變。
骨骼肌肉系統的變化也非常顯著。蝌蚪的肌肉主要分布在尾部,用於游泳推進。隨著變態,這些肌肉被重新分配到四肢和軀幹,以支持跳躍和陸地運動。骨骼系統也從軟骨逐漸轉變為骨質,形成成蛙特有的骨骼結構。
神經系統和感官器官同樣經歷了重大變化。大腦和脊髓進一步發育,以適應更複雜的陸地生活。視覺系統從適應水下環境轉變為適應空氣中的視覺。聽覺系統也發生變化,形成了適應空氣傳播聲音的中耳結構。
值得注意的是,不同種類的蛙類在變態過程中可能表現出顯著的差異。例如,某些樹蛙的變態過程相對較快,可能只需要幾周時間。而某些大型蛙類,如牛蛙(Rana catesbeiana),其變態過程可能持續一年甚至更長時間。這種差異反映了不同蛙類適應特定生態環境的策略。
環境因素對蛙類變態過程有重要影響。例如,食物可用性、水體密度、溫度等因素都可能影響變態的時間和速度。有趣的是,某些蛙類,如墨西哥鐵蛙(Hypopachus variolosus),能夠根據環境條件調整其變態時間。在乾旱或食物匱乏的環境中,這些蛙類可能會加速變態過程,以便盡快離開不利的水環境。
某些蛙類還表現出非典型的變態模式。例如,直接發育型蛙類如所羅門群島的直接發育蛙(Platymantis sp.),其卵直接發育為小蛙,完全跳過了自由生活的蝌蚪階段。這種適應可能是為了應對缺乏適合蝌蚪生存的水體環境。
變態過程中的激素變化不僅影響形態和生理,還可能影響蛙類的行為。例如,隨著變態的進行,許多蛙類開始表現出對陸地環境的偏好,這可能與神經內分泌系統的變化有關。
蛙類的變態過程為我們提供了研究發育生物學和進化生物學的絕佳模型。通過研究蛙類如何在短時間內完成如此劇烈的形態和生理變化,科學家們不僅能夠更好地理解發育過程的調控機制,還可能為人類醫學,如組織再生和器官重建等領域提供新的思路。
此外,蛙類變態過程的敏感性使其成為環境污染的重要指示器。許多環境污染物,特別是內分泌干擾物,可能會影響蛙類的正常變態過程,導致發育異常或死亡。因此,監測蛙類的變態過程成為評估生態系統健康狀況的重要手段之一。
5.4 蛙類的呼吸機制
蛙類的呼吸機制是其生理適應中最為獨特和複雜的部分之一。作為兩棲動物,蛙類必須應對水陸兩種環境的呼吸需求,因此演化出了多樣化且高效的呼吸策略。蛙類的呼吸系統包括肺部呼吸、皮膚呼吸和口腔泵呼吸三種主要機制,這些機制相互配合,使蛙類能夠在不同環境中有效地進行氣體交換。
肺部呼吸是蛙類在陸地環境中的主要呼吸方式。蛙類的肺部結構相對簡單,通常呈現囊狀,內部有少量的肺泡。雖然結構簡單,但蛙類的肺部呼吸效率卻相當高。這主要歸功於其獨特的呼吸機制,稱為鼓式呼吸或口腔泵呼吸(buccal pumping)。由於蛙類缺乏肋骨和橫膈膜,它們無法像哺乳動物那樣通過胸腔擴張和收縮來進行呼吸。相反,蛙類利用口腔底部的肌肉運動來實現空氣的進出。
鼓式呼吸的過程大致如下:首先,蛙類張開鼻孔,降低口腔底部,使空氣通過鼻孔進入口腔。然後,蛙類關閉鼻孔,抬高口腔底部,將空氣推入肺部。呼氣時,肺部的彈性收縮將空氣排出。這個過程可以快速重複,使蛙類能夠有效地更新肺部的空氣。例如,北美豹蛙(Rana pipiens)在安靜時每分鐘可以進行約70次的鼓式呼吸。
值得注意的是,蛙類的鼓式呼吸不僅用於肺部呼吸,還在調節體溫和發聲中發揮重要作用。通過調整鼓式呼吸的頻率和深度,蛙類可以增加或減少蒸發冷卻,從而調節體溫。在發聲時,雄性蛙類利用鼓式呼吸來控制氣流通過聲帶,產生各種複雜的叫聲。
皮膚呼吸是蛙類另一個重要的呼吸機制。蛙類的皮膚薄而多血管,缺乏角質層,這使得氣體能夠直接通過皮膚進行交換。皮膚呼吸對於許多蛙類來說是必不可少的,特別是在水中或高濕度環境中。某些完全水生的蛙類,如非洲爪蟾(Xenopus laevis),甚至可以完全依賴皮膚呼吸生存。
蛙類皮膚呼吸的效率與其皮膚的濕潤度密切相關。為了保持皮膚濕潤,蛙類進化出了多種策略。許多蛙類的皮膚上分佈著大量的黏液腺,能夠分泌黏液保持皮膚濕潤。某些樹蛙,如澳大利亞綠樹蛙(Litoria caerulea),還能分泌特殊的蠟質物質來減少水分蒸發,同時不影響氣體交換。
蛙類的皮膚呼吸還與其循環系統密切相關。蛙類的皮膚下分佈著豐富的畿細血管網,這些血管能夠將氧氣直接輸送到身體各部分。有趣的是,某些蛙類能夠調節皮膚血流量來控制皮膚呼吸的效率。例如,當環境氧氣含量較低時,蛙類可能會增加皮膚血流量以提高氧氣攝取效率。
口腔呼吸是蛙類的第三種呼吸機制。蛙類的口腔和咽部富含血管,能夠直接進行氣體交換。這種呼吸方式在某些情況下特別重要,例如當蛙類需要長時間保持靜止或潛水時。通過口腔呼吸,蛙類可以在不驚動獵物或捕食者的情況下維持基本的氧氣供應。
蛙類的呼吸機制在其生命週期的不同階段會發生顯著變化。蝌蚪階段主要依賴鰓呼吸,隨著變態過程的進行,鰓逐漸退化,肺部和皮膚呼吸系統逐步發育。這個轉變過程是蛙類適應從水生到陸生環境的關鍵。
某些蛙類還發展出了特殊的呼吸適應。例如,某些樹蛙能夠通過鼻腔中的特殊結構進行呼吸,這使得它們能夠在潛入水中時仍然保持呼吸。另一個有趣的例子是肺雨蛙(Barbourula kalimantanensis),這種生活在印度尼西亞加里曼丹島快速流動的溪流中的蛙類完全沒有肺,完全依賴皮膚呼吸。這種極端的適應可能是為了減少浮力,使其能夠更好地在急流中生存。
蛙類的呼吸機制還與其他生理功能密切相關。例如,呼吸過程與酸鹼平衡的維持有關。通過調節呼吸頻率,蛙類可以影響體內二氧化碳的水平,從而調節血液pH值。此外,呼吸過程還與蛙類的發聲機制密切相關。雄性蛙類利用肺部的空氣和特殊的聲囊產生各種複雜的叫聲,這在繁殖季節尤為重要。
蛙類的多樣化呼吸機制使其能夠適應各種環境條件。例如,在乾旱環境中,蛙類可能更多地依賴肺部呼吸,並採取特殊的行為來減少水分流失。而在濕潤或水生環境中,皮膚呼吸可能成為主要的氧氣來源。這種靈活性使得蛙類能夠在從熱帶雨林到乾旱沙漠等各種棲息地中生存。
然而,蛙類的呼吸機制也使其容易受到環境污染的影響。由於皮膚的高度通透性,水體或空氣中的污染物可能直接通過皮膚進入蛙體內。這使得蛙類成為環境污染的敏感指示器,也解釋了為什麼蛙類常常是環境變化的早期受害者。
5.5 蛙類冬眠與夏眠
蛙類的冬眠和夏眠是其適應極端環境條件的重要生理機制。這兩種狀態使蛙類能夠在食物稀缺、溫度極端或環境乾燥的時期存活下來。雖然冬眠和夏眠在表面上看起來相似,但它們在生理機制和觸發因素上存在顯著差異。
冬眠是蛙類為應對寒冷氣候而進化出的生存策略。當環境溫度降低,食物變得稀缺時,許多生活在溫帶和寒帶地區的蛙類會進入冬眠狀態。冬眠過程中,蛙類的新陳代謝率大幅降低,心跳和呼吸頻率顯著減慢。例如,北美木蛙(Rana sylvatica)在冬眠時,其心跳可能從正常時的每分鐘40-50次降至每分鐘僅1-2次。
蛙類的冬眠地點因種類而異。水生性強的蛙類,如綠蛙(Rana clamitans),通常會潛入池塘或湖泊底部的淤泥中冬眠。而一些陸生蛙類,如美洲樹蛙(Hyla versicolor),則可能鑽入土壤深處或樹洞中避寒。有趣的是,某些蛙類還會選擇群體冬眠,這可能有助於減少熱量損失並提高生存率。
蛙類在進入冬眠前會進行一系列生理準備。它們會增加體內的糖原儲備,這些糖原在冬眠期間被轉化為葡萄糖,作為能量來源和天然的「防凍劑」。某些蛙類,如北美木蛙,甚至能夠忍受體液部分結冰的情況。這種極端的耐寒能力得益於它們體內高濃度的葡萄糖,可以將凝固點降低到零下幾度。
夏眠則是蛙類應對高溫乾旱環境的策略。當環境變得炎熱乾燥,水源稀缺時,某些生活在乾旱或半乾旱地區的蛙類會進入夏眠狀態。夏眠期間,蛙類同樣會降低新陳代謝率,減少水分損失,但其生理機制與冬眠有所不同。
澳大利亞的水囊蛙(Cyclorana platycephala)是夏眠適應的典型代表。當乾季來臨時,水囊蛙會鑽入地下,分泌一層特殊的黏液包裹全身,形成一個保護性的茧。這個茧不僅能夠保護蛙類免受乾燥,還能阻擋潛在的捕食者。水囊蛙可以在這種狀態下存活長達幾年之久,直到雨季來臨,地面積水喚醒它們。
非洲牛蛙(Pyxicephalus adspersus)展示了另一種有趣的夏眠策略。這種蛙類會在地下挖掘一個小室,並分泌一種角蛋白質構成的保護層,將自己完全包裹起來。這層保護膜不僅能夠防止水分流失,還允許氣體交換,使蛙類能夠在地下維持最低限度的呼吸。
蛙類在進入夏眠狀態前也會進行生理準備。它們會盡可能多地攝取食物,積累脂肪儲備。同時,它們的新陳代謝會發生改變,開始產生尿素作為主要的代謝產物。尿素在體內積累可以幫助降低水分流失,同時作為一種渦透調節物質,幫助維持細胞的正常功能。
值得注意的是,並非所有的蛙類都會進行典型的冬眠或夏眠。生活在熱帶地區的蛙類可能全年活躍,不需要長期的休眠狀態。而某些生活在溫帶地區的蛙類,如北美豹蛙(Rana pipiens),雖然會在冬季減少活動,但並不進入完全的冬眠狀態。這種策略被稱為「冬眠」(brumation),是爬行動物和兩棲動物特有的一種低活動狀態。
蛙類的冬眠和夏眠狀態可以被環境因素打斷。例如,如果冬季出現異常暖和的天氣,某些蛙類可能會短暫醒來活動。同樣,如果乾季期間出現降雨,夏眠的蛙類可能會被喚醒。這種靈活性使得蛙類能夠最大限度地利用有利的環境條件。
蛙類從冬眠或夏眠中甦醒的過程也是一個複雜的生理過程。它們需要迅速提高新陳代謝率,恢復正常的心跳和呼吸頻率。同時,它們還需要排出在休眠期間積累的代謝產物。對於冬眠的蛙類來說,它們還需要處理體內高濃度的葡萄糖。這個過程通常需要幾個小時到幾天的時間。
蛙類的冬眠和夏眠能力在其生命週期的不同階段可能有所不同。通常,成年蛙類比幼體有更強的耐受能力。這可能是因為成年個體有更多的能量儲備,以及更發達的生理調節機制。
氣候變化對蛙類的冬眠和夏眠行為產生了顯著影響。全球變暖可能導致某些地區的冬季變短或變暖,這可能影響蛙類的冬眠時機和持續時間。同樣,極端天氣事件的增加,如長期乾旱,可能迫使更多蛙類進入夏眠狀態,或延長夏眠時間。這些變化可能對蛙類的生存和繁殖產生深遠影響。
研究蛙類的冬眠和夏眠機制不僅有助於我們理解這些生物的生存策略,還可能為人類醫學帶來啟發。例如,了解蛙類如何在體液部分結冰的情況下生存,可能為開發新的器官保存技術提供思路。同樣,研究蛙類如何在長期缺水的情況下維持生理功能,可能為治療某些人類疾病提供新的思路。
6. 蛙類行為與溝通
蛙類的行為與溝通是一個豐富多彩的研究領域,展現了這些兩棲動物的複雜性和適應能力。在蛙類的世界中,聲音是最主要的交流方式。雄蛙通過發出獨特的叫聲來吸引雌蛙、宣示領地或警告同類。這些聲音不僅在頻率和音調上各不相同,還能傳達特定的信息。例如,某些樹蛙物種能夠通過改變其叫聲的音調來表達不同的訊息,如求偶或警戒。
除了聲音,視覺信號也在蛙類的溝通中扮演重要角色。許多蛙物種具有鮮艷的體色或特殊的斑紋,這些不僅用於偽裝,還可作為吸引異性或警告捕食者的信號。一些樹蛙甚至能夠迅速改變體色來適應不同的情況。
社會行為在蛙類中也十分普遍。某些物種會形成繁殖聚集,雄蛙們在水域邊緣排成一列,爭相吸引雌蛙的注意。領域行為是另一個有趣的方面,特別是在繁殖季節,雄蛙會積極地保衛自己的區域,通過叫聲或身體接觸來驅趕入侵者。
人類對蛙鳴的解讀和研究也是一個引人入勝的話題。科學家們通過分析蛙鳴的頻率、持續時間和模式,不僅能識別不同的物種,還能評估種群的健康狀況和環境變化。這種研究對於生態監測和保育工作具有重要意義,使我們能更好地理解和保護這些獨特的生物。
6.1 蛙類發聲與聲學交流
蛙類的發聲與聲學交流是一個引人入勝的研究領域,展現了這些兩棲動物驚人的多樣性和適應能力。在蛙類的世界中,聲音不僅是一種簡單的交流方式,更是一種複雜的生存策略和演化成果。
蛙類發聲的核心機制涉及一個稱為鳴囊的特殊結構。這是一個位於喉部或口角的薄膜囊,可以迅速膨脹和收縮。當蛙類將空氣從肺部擠壓到鳴囊時,就會產生聲音。不同物種的鳴囊大小、形狀和位置各不相同,這直接影響了它們所能發出的聲音類型。
蛙類的叫聲在頻率、音調、節奏和複雜性上都有巨大的變化。有些物種發出簡單的單音調鳴叫,而其他物種則能產生複雜的多音節叫聲。例如,台灣特有種莫氏樹蛙(Rhacophorus moltrechti)的叫聲就像是一連串的「嘰-嘰-嘰」,而面天樹蛙(Kurixalus idiootocus)的叫聲則更像是「咯-咯-咯」的連續音。
蛙類發聲的主要目的之一是求偶。雄蛙通過發出獨特的叫聲來吸引雌蛙。這些叫聲不僅傳達了物種的身份,還可能包含個體的特質信息,如體型、健康狀況或基因品質。研究發現,某些物種的雌蛙能夠根據雄蛙叫聲的特定特徵來選擇配偶。例如,在某些樹蛙物種中,雌蛙偏好頻率較低的叫聲,因為這通常表示雄蛙體型較大或年齡較長,可能擁有更好的基因。
除了求偶,蛙類的叫聲還有其他重要功能。領域宣示是其中之一,雄蛙通過發聲來標記和維護自己的領地。當其他雄蛙靠近時,領地擁有者會發出警告性的叫聲。如果入侵者不退卻,可能會導致聲音對峙,有時甚至演變成身體衝突。
警戒叫聲是另一種重要的聲音交流方式。當察覺到危險時,某些蛙類會發出特殊的警戒聲,提醒同類注意潛在的威脅。這種行為不僅保護了個體,也增加了種群的整體生存機會。
蛙類的聲學交流還展現了令人驚嘆的適應性。在嘈雜的環境中,如瀑布附近或熱鬧的蛙類合唱中,某些物種已經演化出特殊的策略來確保其信息能被有效傳遞。例如,有些蛙類會在其他蛙類叫聲的間隙發聲,以避免信號被掩蓋。還有一些物種能夠調整其叫聲的頻率或音量,以適應不同的聲學環境。
蛙類的發聲行為還受到多種環境因素的影響。溫度、濕度、季節甚至月相都可能影響蛙類的叫聲特徵和發聲頻率。例如,在溫暖的夜晚,蛙類通常更為活躍,叫聲也更頻繁。這種對環境條件的敏感性使得蛙類成為生態變化的優秀指示器。
有趣的是,某些蛙類已經演化出非聲音的交流方式,特別是在噪音較大的環境中。例如,一些生活在湍急溪流旁的蛙類物種會使用視覺信號,如揮動腳掌或膨脹喉部,來吸引異性或警告同類。這種行為適應展示了演化的創造力和蛙類的適應能力。
人類對蛙類聲音的研究和應用也是一個迷人的領域。科學家們使用高科技錄音設備和聲譜分析軟件來研究蛙鳴,這不僅有助於物種識別和種群監測,還能幫助我們了解環境變化對蛙類的影響。例如,通過長期監測某一地區的蛙鳴,研究人員可以追蹤物種組成的變化,評估棲息地質量,甚至預測某些環境問題。
此外,蛙類的發聲機制也啟發了一些技術創新。例如,工程師們研究蛙類如何在水下發聲,以開發更好的水下通訊設備。蛙類獨特的發聲方式也為音樂家和聲音藝術家提供了靈感,創造出融合自然音色的新型音樂作品。
然而,人類活動對蛙類的聲學交流造成了一些負面影響。噪音污染、棲息地破壞和氣候變化都可能干擾蛙類的正常發聲和交流。例如,城市噪音可能掩蓋蛙類的叫聲,減少它們成功繁殖的機會。這提醒我們需要在發展過程中考慮到野生動物的需求,採取措施減少人類活動對自然生態系統的干擾。
蛙類的發聲與聲學交流不僅是一個豐富的科學研究領域,也是自然界中一個迷人的聲音景觀。從單調的「呱呱」聲到複雜的旋律性鳴叫,蛙類用它們的聲音譜寫著地球上最古老和最動聽的自然交響曲之一。理解和欣賞這種交流方式,不僅能增進我們對這些兩棲動物的認識,還能加深我們對整個生態系統複雜性和脆弱性的理解。
6.2 蛙類視覺信號
蛙類的視覺信號是一個引人入勝的研究領域,展示了這些兩棲動物在溝通和適應方面的驚人能力。儘管蛙類以其鳴叫聲而聞名,但它們的視覺信號同樣複雜多樣,在種內和種間交流中扮演著重要角色。
蛙類的視覺信號主要通過體色、體型和行為模式來表現。體色是最顯著的視覺特徵之一,不同物種呈現出豐富多彩的色彩組合。這些色彩不僅有助於偽裝,還可作為警告信號或求偶展示。例如,箭毒蛙(Dendrobatidae)家族以其鮮艷的體色而聞名,這些明亮的紅色、黃色或藍色不僅美麗,還向潛在捕食者傳達了毒性警告。相反,一些樹蛙物種則擁有綠色或褐色的保護色,能完美融入周圍環境。
有趣的是,某些蛙類物種能夠根據環境或生理狀態改變體色。這種現象稱為生理性變色,是一種重要的視覺信號機制。例如,一些樹蛙能在數分鐘內從亮綠色變為深褐色,以適應不同的背景或光照條件。這種能力不僅有助於躲避捕食者,還可能在種內交流中發揮作用,如表達壓力或準備繁殖的狀態。
蛙類的體型和姿態也是重要的視覺信號。在求偶季節,許多物種的雄性會採取特定的姿勢來吸引雌性或威嚇競爭對手。例如,某些樹蛙物種的雄性會膨脹喉部,形成一個明顯的聲囊,不僅用於發聲,還作為視覺展示。有些地棲蛙類則會抬高身體,展示胸部或腹部的鮮艷色彩,這種行為既是求偶信號,也可能是對其他雄性的威脅展示。
蛙類的眼睛也是視覺信號的重要組成部分。許多物種擁有大而明亮的眼睛,不同的虹膜顏色和圖案可能傳達物種特異性信息。一些研究表明,眼睛的大小和顏色可能與個體的健康狀況和吸引力相關,影響配偶選擇。
在某些蛙類物種中,腳部的顏色和圖案也是重要的視覺信號。例如,一些生活在湍急溪流中的蛙類會有鮮艷的腳蹼,當它們在水中游動時,這些顏色會引起注意,可能用於種內識別或求偶展示。
蛙類還通過特定的行為模式來傳遞視覺信號。例如,許多物種的雄性會進行所謂的「踏腳舞」,即快速抬起和放下後腿,這種行為既可以吸引雌性,也可能用於驅趕入侵者。另一種常見的視覺行為是「點頭」,蛙類通過快速上下移動頭部來傳達各種信息,如領域宣示或求偶意圖。
在一些噪音較大的環境中,如瀑布附近或熱鬧的蛙類聚集地,視覺信號變得尤為重要。在這些情況下,聲音可能被環境噪音掩蓋,因此一些蛙類物種演化出更依賴視覺信號的交流方式。例如,某些生活在山溪旁的蛙類會通過誇張的腳部動作或身體姿態來吸引異性,這種適應展示了蛙類在不同環境中的交流靈活性。
蛙類的視覺信號還與其生活環境密切相關。樹棲物種通常擁有更鮮艷的顏色和更複雜的視覺展示行為,這可能與它們在樹冠層中需要更顯眼的信號有關。相比之下,地棲或水生物種的視覺信號可能更加微妙,更多地依賴於姿態和行為而非顏色。
值得注意的是,蛙類的視覺信號並非孤立存在,而是與其他感官信號緊密結合。例如,許多物種的雄性在發出求偶鳴叫的同時,會膨脹喉囊或採取特定姿勢,形成一個多感官的信號組合。這種綜合信號可能比單一的聲音或視覺信號更有效地傳達信息。
蛙類視覺信號的研究對於理解它們的行為生態學和演化生物學具有重要意義。通過研究不同物種的視覺信號,科學家們可以了解環境因素如何塑造交流策略,以及這些信號如何影響種群動態和物種形成過程。例如,某些近緣物種可能主要通過視覺信號的差異來維持物種界限,防止雜交。
此外,蛙類的視覺信號研究還為生態保護提供了重要信息。由於許多蛙類物種對環境變化非常敏感,它們的視覺信號可能會因環境壓力而改變。例如,棲息地破壞或污染可能導致某些物種的體色變得暗淡,或改變其正常的行為模式。因此,監測蛙類的視覺信號可以作為評估生態系統健康的一個重要指標。
人類活動對蛙類視覺信號的影響也是一個值得關注的問題。光污染可能干擾夜行性蛙類的正常視覺交流,而棲息地改變可能影響背景與蛙類體色的對比度,降低視覺信號的有效性。理解這些影響對於制定有效的保護策略至關重要。
蛙類的視覺信號不僅是科學研究的對象,也是自然愛好者和攝影師的靈感來源。這些豐富多彩、形態各異的生物為我們展示了大自然的創造力和多樣性。通過觀察和記錄蛙類的視覺信號,我們不僅能增進對這些迷人生物的了解,還能加深對生態系統複雜性的認識。
6.3 蛙類社會行為
蛙類的社會行為是一個複雜而迷人的的研究領域,展現了這些兩棲動物令人驚訝的社交能力和行為多樣性。儘管蛙類通常被認為是相對獨立的生物,但許多物種實際上具有豐富的社會互動和組織結構。
繁殖季節是蛙類社會行為最為活躍和複雜的時期。在這個時候,許多物種會形成大規模的繁殖聚集,這種現象被稱為「蛙類合唱團」。在這些聚集中,雄性蛙類會聚集在水域附近,發出響亮的求偶鳴叫。這種行為不僅吸引雌性,也形成了一種臨時的社會結構。例如,在某些樹蛙物種中,雄性會在樹枝或葉子上排成一列,形成所謂的「合唱線」。這種排列不是隨機的,而是遵循一定的社會等級制度,其中較大或更有經驗的雄性通常佔據更有利的位置。
在這些繁殖聚集中,蛙類展現出複雜的社會互動。雄性之間可能會發生競爭和衝突,包括聲音對抗和身體接觸。有趣的是,某些物種已經演化出降低衝突的策略。例如,有些蛙類會通過改變叫聲的音調或節奏來表明自己的體型或競爭能力,從而避免不必要的身體衝突。
某些蛙類物種還表現出令人驚訝的協作行為。例如,南美的負子蛙(Ates obstetricans)展示了雄性之間的合作育幼行為。在這個物種中,多個雄性會共同照顾一窩卵,輪流將卵串纏繞在後腿上並保持濕潤,直到它們孵化。這種行為不僅增加了後代的存活機會,還形成了一種獨特的社會結構。
蛙類的領域行為也是其社會行為的重要組成部分。許多物種的雄性會建立和守衛自己的領地,特別是在繁殖季節。這種行為涉及複雜的社會互動,包括聲音警告、視覺展示和身體接觸。有趣的是,某些物種的領域大小和品質與個體的社會地位密切相關。優勢個體通常能夠佔據和守衛更大、更優質的領地。
值得注意的是,某些蛙類物種表現出類似於哺乳動物的社會行為。例如,委內瑞拉的利特爾樹蛙(Dendropsophus ebraccatus)被發現具有類似家庭結構的社會組織。在這個物種中,成對的雄性和雌性會長期共同生活,共同防衛領地和照顧後代。這種行為挑戰了我們對兩棲動物社會結構的傳統理解。
蛙類的社會行為還包括集體防禦機制。某些物種在面對捕食者時會採取群體行動。例如,一些樹蛙物種會在感知到威脅時集體發出警戒叫聲,這種行為不僅保護了個體,還增強了整個群體的生存機會。
在非繁殖季節,許多蛙類物種也表現出有趣的社會行為。例如,某些樹蛙物種會形成小型的社交群體,共同棲息在樹洞或葉片下。這種行為可能有助於調節體溫、減少水分損失,並提供額外的保護。
蛙類的社會學習也是一個新興的研究領域。儘管長期以來認為兩棲動物缺乏複雜的學習能力,但最近的研究表明,某些蛙類物種能夠通過觀察同類來學習新的行為。例如,有研究發現,某些樹蛙能夠通過觀察其他個體的跳躍行為來學習避開危險。
蛙類的親代照顧行為是其社會行為中特別引人注目的一面。雖然大多數蛙類並不提供直接的親代照顧,但有些物種表現出令人驚訝的育幼行為。例如,毒箭蛙(Dendrobates)家族的一些物種會將蝌蚪背負到安全的水域。在這個過程中,成體不僅要定期為蝌蚪提供營養,還要保護它們免受捕食者的侵害。這種行為不僅增加了後代的存活率,還形成了一種獨特的親子社會關係。
蛙類的求偶行為也展現了複雜的社會互動。除了常見的鳴叫和視覺展示,某些物種還發展出了獨特的求偶儀式。例如,有些樹蛙物種的雄性會進行所謂的「蛙式舞蹈」,這種行為包括特定的身體動作和節奏,用以吸引雌性的注意。這種複雜的求偶行為不僅是一種社會互動,還可能是雌性評估潛在配偶品質的一種方式。
蛙類的群體動態也是社會行為研究的重要方面。在某些物種中,個體之間存在明顯的等級制度。這種等級結構可能基於體型、年齡或經驗,並影響個體在繁殖、覓食和棲息地選擇等方面的機會。了解這些社會結構對於理解蛙類種群動態和演化過程至關重要。
值得一提的是,蛙類的社會行為還受到環境因素的強烈影響。例如,棲息地破碎化可能改變蛙類的社會互動模式,影響它們的繁殖成功率和種群結構。同樣,氣候變化也可能影響蛙類的活動模式和社會行為,進而影響整個生態系統的平衡。
人類活動對蛙類社會行為的影響也是一個值得關注的問題。例如,光污染可能干擾夜行性蛙類的正常社交活動,而噪音污染可能影響它們的聲音交流。了解這些影響對於制定有效的保護策略至關重要。
研究蛙類的社會行為不僅有助於我們更好地理解這些迷人的生物,還為我們提供了洞察生態系統複雜性的窗口。通過觀察和分析蛙類的社會互動,我們可以更深入地理解生物多樣性的價值和生態系統的脆弱性。這些知識對於制定有效的保護策略和管理計劃至關重要,尤其是在面對全球環境變化的背景下。
6.4 蛙類領域行為
蛙類的領域行為是一個複雜而引人入勝的研究領域,展現了這些兩棲動物在生態適應和社會互動方面的獨特性。領域行為在蛙類中普遍存在,但其表現形式和強度因物種、性別、季節和環境條件而異。
領域行為最常見於雄性蛙類,特別是在繁殖季節。這種行為的主要目的是確保獲得和控制優質的繁殖資源,如適合產卵的水域或有利的鳴叫位置。通過建立和維護領地,雄性蛙類可以增加吸引雌性和成功繁殖的機會。
蛙類建立領地的方式多種多樣。聲音是最常見且最有效的領域宣示方式。雄性蛙類會發出特定的領域叫聲,這種叫聲不僅用於吸引雌性,也向其他雄性傳達「此地已有主人」的信息。例如,在台灣常見的盤古蟾蜍(Bufo bankorensis)中,雄性會發出持續的低沉叫聲來標記其領地。這種叫聲的頻率和強度可能會根據潛在入侵者的接近程度而改變。
視覺信號也在蛙類的領域行為中扮演重要角色。許多物種會採取特定的姿勢或展示行為來宣示領地。例如,某些樹蛙物種會膨脹喉囊或採取高姿態的身體姿勢來增加自身的視覺體積,從而顯得更具威脅性。這種行為不僅用於威嚇其他雄性,也可能是向雌性展示自身實力的一種方式。
物理接觸是蛙類領域行為中最直接但也最具風險的形式。當聲音和視覺警告無效時,雄性蛙類可能會發生身體衝突。這種衝突可能包括推擠、抓撓,甚至是摔跤式的搏鬥。例如,在某些大型蛙類物種中,雄性間的領域衝突可能會演變成激烈的身體接觸,有時甚至導致輕微的傷害。
蛙類的領域大小和品質通常與個體的特徵密切相關。較大、更健康或更有經驗的雄性往往能夠獲得和維護更大、更優質的領地。這種領地質量的差異直接影響繁殖成功率,因為優質領地通常意味著更好的產卵場所或更有利的鳴叫位置。
有趣的是,某些蛙類物種展現出靈活的領域策略。例如,在資源豐富的地區,領地可能相對較小且密集;而在資源稀缺的地區,領地可能更大且分散。這種適應性策略允許蛙類根據環境條件優化其繁殖機會。
蛙類的領域行為還受到時間因素的影響。在許多物種中,領域行為主要集中在繁殖季節。然而,某些物種可能全年維護領地,特別是那些棲息在穩定環境中的物種。例如,某些生活在熱帶雨林中的樹蛙可能全年都在維護其領地,這些領地不僅用於繁殖,也作為重要的覓食和休息區域。
值得注意的是,雖然領域行為在雄性中最為常見,但某些物種的雌性也表現出領域行為。這種情況通常出現在資源有限或繁殖機會稀缺的環境中。例如,在某些高山湖泊中生活的蛙類物種,由於適合產卵的地點有限,雌性可能會為這些稀缺資源而競爭並建立臨時領地。
蛙類的領域行為還涉及複雜的決策過程。個體需要權衡維護領地的成本和收益。守衛領地需要消耗能量,可能增加被捕食的風險,但同時也提供了繁殖和資源獲取的機會。因此,蛙類已經演化出精細的機制來評估威脅程度並做出相應反應。例如,某些物種能夠通過入侵者的叫聲來判斷其體型和威脅程度,並據此決定是否進行積極防禦。
人類活動對蛙類的領域行為產生了重大影響。棲息地破碎化和污染可能改變可用的領地資源,迫使蛙類改變其領域行為模式。例如,在城市化地區,適合繁殖的水域可能變得稀缺,導致領域競爭加劇。此外,噪音污染可能干擾蛙類的聲音通訊,影響其維護領地的能力。
氣候變化也對蛙類的領域行為產生深遠影響。溫度和降水模式的改變可能影響蛙類的活動週期和繁殖時間,進而影響其領域行為的表現。例如,在某些地區,由於氣候變暖,繁殖季節可能提前或延長,改變了傳統的領域建立和維護模式。
研究蛙類的領域行為不僅有助於我們理解這些迷人的生物的生態學和行為學,還為保護工作提供了重要見解。通過了解蛙類如何選擇和維護領地,保護生物學家可以更好地設計和管理保護區,確保為不同物種提供適當的棲息環境。
此外,蛙類的領域行為研究還為我們提供了研究動物社會行為演化的寶貴模型。由於蛙類在演化樹上的位置,研究它們的行為可以幫助我們理解更複雜社會行為的起源和發展。
蛙類領域行為的研究方法也在不斷進步。除了傳統的觀察和記錄方法,研究人員現在還使用先進的技術,如聲音分析軟件、微型GPS追蹤器和紅外相機,來詳細記錄和分析蛙類的領域行為。這些技術使我們能夠更深入地了解蛙類如何在時間和空間上使用其領地。
領域行為在蛙類的生活史中扮演著核心角色,影響其繁殖成功、資源獲取和種群動態。隨著我們對這一行為的理解不斷深入,我們不僅能更好地欣賞這些兩棲動物的複雜性,還能為其保護提供更有力的科學依據。在面對全球環境變化的背景下,這些知識對於制定有效的保護策略和管理計劃至關重要。
6.5 蛙鳴與人類解讀
蛙鳴是大自然中最引人注目的聲音之一,長期以來一直吸引著人類的興趣和好奇。蛙類發出的各種聲音不僅是它們之間交流的重要方式,也為人類提供了豐富的生態信息和文化意涵。隨著科技的進步和研究方法的改進,人類對蛙鳴的解讀和理解也在不斷深化,揭示了這些兩棲動物聲音世界的複雜性和多樣性。
蛙鳴的主要功能是用於種內交流,特別是在求偶和領域宣示方面。每個物種都有其獨特的叫聲,這使得科學家能夠通過聲音來識別不同的蛙類。例如,台灣特有種的莫氏樹蛙(Rhacophorus moltrechti)發出的是一種類似「嘰-嘰-嘰」的連續短促叫聲,而盤古蟾蜍(Bufo bankorensis)則發出持續的低沉鳴叫。這些獨特的聲音特徵不僅幫助蛙類識別同類,也為人類研究者提供了寶貴的識別工具。
人類對蛙鳴的解讀經歷了從直觀感受到科學分析的過程。在早期,人們主要依靠耳朵和經驗來識別不同的蛙類叫聲。隨著錄音技術的發展,科學家們開始系統地記錄和分析蛙鳴。現代聲譜分析技術的應用,使得研究人員能夠精確地測量蛙鳴的各種聲學參數,如頻率、音調、節奏和持續時間。這些數據不僅用於物種識別,還能揭示蛙類個體的特徵,如體型、年齡和健康狀況。
蛙鳴分析在生態監測中扮演著重要角色。通過長期記錄和分析特定區域的蛙鳴,科學家們可以追蹤蛙類種群的變化趨勢,評估棲息地質量,甚至預測環境變化。例如,某些蛙類對環境污染特別敏感,它們叫聲的變化或消失可能是水質惡化的早期警示信號。在氣候變化研究中,蛙類的繁殖叫聲時間可以作為季節變化的指標,幫助科學家們追蹤全球變暖對生態系統的影響。
人類對蛙鳴的解讀還涉及文化和民俗方面。在許多文化中,蛙鳴被視為天氣變化的預兆。例如,在台灣的民間傳說中,蛙類大量鳴叫常被解讀為即將下雨的徵兆。雖然這種解讀可能不完全準確,但反映了人類長期觀察自然的結果,也體現了蛙類在人類文化中的重要地位。
隨著城市化的推進,蛙鳴在某些地區變得越來越稀少。這種變化引發了人們對聲景(soundscape)保護的關注。聲景生態學家們開始研究如何保護和恢復自然聲音環境,其中蛙鳴是重要的組成部分。在一些地區,人們甚至開展了「蛙鳴導覽」活動,旨在提高公眾對這些聲音及其生態價值的認識。
近年來,人工智能和機器學習技術的應用為蛙鳴研究帶來了新的機遇。研究人員開發了能夠自動識別和分類蛙鳴的算法,大大提高了數據處理的效率和準確性。這些技術使得大規模、長期的蛙類聲音監測成為可能,為生態研究和保護工作提供了強有力的工具。
蛙鳴的研究還涉及聲音傳播和感知的物理學原理。科學家們發現,不同物種的蛙類已經進化出適應特定環境的發聲和聽覺機制。例如,生活在湍急溪流旁的蛙類往往具有更高頻率的叫聲,以便在嘈雜的背景中傳播。理解這些適應性特徵有助於我們更好地解讀蛙鳴在不同生態環境中的功能和意義。
人類對蛙鳴的解讀還延伸到了行為生態學領域。通過分析蛙鳴的細微變化,研究人員可以推斷出蛙類的情緒狀態和社會互動。例如,某些物種在面對捕食者時會發出特殊的警戒叫聲,而在競爭激烈的繁殖季節,雄性蛙類的叫聲可能會變得更加頻繁和響亮。
蛙鳴研究還為生物聲學技術的發展提供了靈感。工程師們研究蛙類如何在嘈雜的環境中有效傳遞聲音信息,並將這些原理應用於改進人類的通信技術。例如,某些水下通信系統的設計就借鑒了蛙類在水中傳播聲音的機制。
在教育領域,蛙鳴為環境教育提供了生動的教學素材。許多學校和自然教育中心開展「夜間蛙類觀察」活動,讓學生通過聆聽和識別蛙鳴來了解生物多樣性和生態系統。這種直接的聽覺體驗往往能激發學生對自然科學的興趣,培養他們的生態意識。
然而,人類活動對蛙類的聲音交流產生了顯著影響。噪音污染、棲息地破壞和氣候變化都可能干擾蛙類的正常鳴叫行為。研究表明,在城市環境中,蛙類可能不得不改變其叫聲的頻率或強度以適應背景噪音。這種適應雖然有助於蛙類在短期內維持交流,但可能對其長期生存和繁衍產生不利影響。
蛙鳴研究也為生物多樣性保護提供了重要依據。通過分析特定地區的蛙鳴多樣性,科學家們可以評估該地區的生態健康狀況。在一些生物多樣性熱點地區,蛙鳴監測已經成為評估保護效果的重要指標之一。

7. 蛙類繁殖與生活史
蛙類的繁殖與生活史展現了自然界中最為迷人的的生物過程之一。這些兩棲動物採用多樣化的繁殖策略,適應不同的環境條件。大多數蛙類採用外部受精,雌性在水中產卵,雄性同時釋放精子。然而,一些物種發展出獨特的繁殖方式,如樹蛙將卵產在樹葉上,或負子蛙將卵攜帶在背上。
蛙類的生活史通常包括卵、蝌蚪和成體三個階段。每個階段都面臨不同的生存挑戰和選擇壓力。卵期是最脆弱的階段,容易受到捕食和環境因素的影響。蝌蚪階段經歷戲劇性的的形態變化,從水生到適應陸地生活。這個變態過程涉及複雜的生理和形態學變化,包括發展肺部、吸收尾部和改變消化系統。
某些蛙類物種表現出特殊的親代照顧行為。例如,毒箭蛙會將蝌蚪背負到安全的水域,並定期餵食未受精的卵。這種行為大大提高了後代的存活率。蛙類的繁殖季節和生活史時機往往與環境條件密切相關,如溫度、降雨和食物可用性。了解這些繁殖和生活史策略對於蛙類保護和生態系統管理至關重要,尤其是在面對棲息地破壞和氣候變化的背景下。
7.1 蛙類交配系統
蛙類的交配系統展現了自然界中令人驚嘆的多樣性和適應性,反映了這些兩棲動物在不同環境中繁衍後代的策略。蛙類的交配系統主要可分為幾種類型,每種類型都有其獨特的特徵和生態意義。
最常見的蛙類交配系統是爆發式繁殖。在這種系統中,大量的雄性和雌性在短時間內聚集在特定的繁殖地點,通常是在雨季開始或特定的環境條件出現時。這種繁殖方式在許多池塘繁殖的蛙類中很常見,如台灣的盤古蟾蜍(Bufo bankorensis)。在爆發式繁殖中,競爭非常激烈,雄性通常會試圖抱住任何接近的個體,有時甚至會形成多雄抱對(multiple amplexus)的情況。這種繁殖策略雖然看似混亂,但能確保大量的卵同時受精,從而增加種群的整體繁殖成功率。
另一種常見的交配系統是延長型繁殖。在這種系統中,繁殖季節持續較長時間,可能延續數週甚至數月。這種繁殖方式通常見於樹蛙和一些生活在穩定環境中的蛙類物種。例如,台灣特有種莫氏樹蛙(Rhacophorus moltrechti)就採用這種繁殖策略。在延長型繁殖系統中,雄性通常會建立和防衛領地,通過鳴叫吸引雌性。這種系統允許雌性有更多時間和機會選擇理想的配偶,從而可能提高後代的質量。
某些蛙類物種表現出一夫多妻制(pogyny)的交配系統。在這種系統中,少數優勢雄性會與多個雌性交配。這種策略通常出現在資源分佈不均或雄性之間存在顯著質量差異的情況下。例如,某些大型蛙類物種中,體型較大或佔據更優質領地的雄性往往能吸引更多雌性。
相對罕見但同樣有趣的是一妻多夫制(poandry)交配系統。在這種系統中,一個雌性會與多個雄性交配。這種策略可能出現在雌性稀少或環境條件不穩定的情況下。例如,某些生活在乾旱地區的蛙類物種中,雌性可能會與多個雄性交配以確保受精成功率。
一些蛙類物種展現出近似於一夫一妻制(monogamy)的行為。雖然真正的終生一夫一妻制在蛙類中極為罕見,但某些物種會在一個繁殖季節內保持配對關係。這種行為通常與特殊的繁殖需求或親代照顧有關。例如,某些樹蛙物種會共同選擇和準備產卵場所,並在整個繁殖季節中保持親密關係。
有趣的是,某些蛙類物種表現出混合型的交配系統。例如,在同一個種群中,可能同時存在爆發式繁殖和延長型繁殖的個體。這種策略的多樣性可能是對不同環境條件和機遇的適應。
蛙類的交配系統還涉及複雜的求偶行為。在許多物種中,雄性通過發出特定的求偶鳴叫來吸引雌性。這些叫聲不僅用於物種識別,還可能傳達個體的質量信息。例如,某些樹蛙物種的雌性能夠根據雄性叫聲的頻率和持續時間來評估其體型和體能狀況。
除了聲音,視覺信號在某些蛙類的交配系統中也扮演重要角色。一些物種的雄性會展示鮮艷的體色或特殊的身體姿態來吸引雌性。例如,某些樹蛙物種的雄性會膨脹喉囊或展示鮮艷的側面體色來增加自身的吸引力。
在某些蛙類物種中,化學信號也參與交配系統。雄性可能會釋放特定的信息素來吸引雌性或標記領地。這種交流方式在水生環境中特別有效,可以在視覺和聽覺受限的情況下傳遞信息。
蛙類的交配系統還受到環境因素的強烈影響。例如,在資源豐富的環境中,可能出現更多的一夫多妻制或延長型繁殖系統。而在資源稀缺或環境不穩定的地區,爆發式繁殖可能更為普遍。氣候變化和棲息地破壞等因素也可能導致蛙類交配系統的改變,進而影響種群的繁殖成功率和長期生存。
研究蛙類的交配系統不僅有助於我們理解這些迷人的生物的生態和演化,還為保護工作提供了重要的科學基礎。通過了解不同物種的繁殖需求和策略,保護生物學家可以更好地設計和管理保護區,確保為不同的繁殖系統提供適當的環境條件。
此外,蛙類交配系統的研究還為我們提供了理解動物行為演化的寶貴見解。通過比較不同物種和種群的交配策略,我們可以探索環境因素、遺傳基礎和演化壓力如何塑造動物的繁殖行為。
隨著研究技術的進步,如基因分析和遠程追蹤,科學家們能夠更深入地研究蛙類的交配系統。這些新方法使我們能夠精確地確定親子關係,追蹤個體的繁殖歷史,並在更大的時空尺度上理解蛙類的繁殖動態。
7.2 蛙類繁殖策略
蛙類的繁殖策略展現了自然界中令人驚嘆的多樣性和創新性,反映了這些兩棲動物在數百萬年演化過程中對不同環境的適應。這些策略涵蓋了從卵的產生到後代的照顧等各個方面,每種策略都有其獨特的生態意義和演化背景。
最常見的蛙類繁殖策略是水生卵繁殖。在這種策略中,雌蛙在水中產下大量的卵,通常以卵團或卵串的形式。這些卵被一種特殊的凝膠物質包裹,既能保護卵不被捕食,又能保持適當的濕度。例如,台灣常見的盤古蟾蜍(Bufo bankorensis)就採用這種策略,在淺水區產下長長的卵串。這種大量產卵的策略雖然看似簡單,但實際上是對高死亡率的一種適應。由於卵和蝌蚪面臨著來自捕食者和環境的多種威脅,大量產卵能確保至少有一部分後代存活下來。
與水生卵策略相對的是陸生卵繁殖策略。某些蛙類物種選擇將卵產在陸地上,通常是在潮濕的環境中,如苔蘚、落葉或樹洞中。這種策略的優點是可以避免水中的許多捕食者,但也面臨著乾燥的風險。採用這種策略的蛙類通常會有特殊的適應,如產生更具彈性的卵殼或選擇特別潮濕的微環境。例如,某些樹蛙物種會將卵產在懸掛在水面上的葉子上,既避免了水中捕食者,又確保了蝌蚪孵化後能夠直接掉入水中。
泡沫巢繁殖是另一種獨特的策略。在這種策略中,蛙類會在水邊或樹上製造一個由泡沫組成的巢。雌蛙在產卵過程中會分泌一種特殊的蛋白質,當這種蛋白質與空氣接觸並被攪拌時,就會形成泡沫。卵被包裹在這些泡沫中,既能得到保護,又能保持濕潤。這種策略特別適合在乾旱或半乾旱地區繁殖的蛙類。泡沫巢不僅能保護卵免受捕食和乾燥,還能提供一個穩定的溫度環境。
一些蛙類物種採用了背負式繁殖策略。在這種策略中,成體(通常是雄性)會將受精卵或發育中的蝌蚪背在背上。這種策略提供了極高程度的親代保護,大大提高了後代的存活率。例如,負子蛙科(Atidae)的成員就以這種方式照顧後代。雄性負子蛙會將卵串纏繞在後腿上,並保持卵的濕潤,直到蝌蚪孵化並能夠獨立生活。
有些蛙類發展出了更為極端的直接發育策略。在這種策略中,整個蝌蚪階段都在卵內完成,孵化出來的就是小青蛙。這種策略避免了脆弱的水生蝌蚪階段,特別適合生活在缺乏穩定水體的環境中的蛙類。例如,某些生活在熱帶雨林地面的蛙類物種就採用這種策略,它們的卵直接發育成小蛙,完全跳過了游泳蝌蚪的階段。
一些蛙類還展現出複雜的育兒行為。例如,某些樹蛙物種的雌性會選擇特定的樹洞或葉片卷起的空間作為「育兒室」。牠們會定期回來檢查卵的狀況,並在必要時添加水分。更令人驚奇的是,某些物種如毒箭蛙,會將孵化的蝌蚪背負到不同的小水體中,並定期回來餵食未受精的卵,這種行為極大地提高了後代的存活率。
蛙類的繁殖策略還包括時間上的調節。許多物種會根據環境條件來決定繁殖的時機。例如,在乾旱地區生活的蛙類可能會等待雨季的到來才開始繁殖。這種策略確保了後代有足夠的水資源來完成發育。相反,在熱帶雨林中,某些蛙類可能全年都有繁殖活動,但會在食物最豐富的季節加強繁殖力度。
值得注意的是,某些蛙類物種表現出繁殖策略的靈活性。牠們可能根據當前的環境條件來調整策略。例如,在水資源豐富的年份,可能會採用水生卵策略;而在乾旱的年份,則可能轉向更保守的策略,如減少產卵量或選擇更安全的產卵地點。
蛙類的繁殖策略還涉及性別角色的分配。在大多數物種中,雄性負責吸引配偶和防衛領地,而雌性則負責選擇配偶和決定產卵位置。然而,在某些物種中,這種角色可能會發生逆轉。例如,在某些稀有的情況下,雌性可能會主動競爭交配機會,而雄性則更多地參與後代照顧。
研究蛙類的繁殖策略不僅有助於我們理解這些迷人的生物的生態和演化,還為保護工作提供了重要的指導。通過了解不同物種的繁殖需求和策略,保護生物學家可以更好地設計和管理保護區,確保為不同的繁殖策略提供適當的環境條件。例如,對於採用樹洞繁殖策略的樹蛙,保護老齡樹木可能是關鍵;而對於需要臨時水體的物種,維護小型濕地的多樣性則至關重要。
7.3 蛙類幼體發育
蛙類幼體發育是一個令人著迷的生物學過程,展示了生命形態的驚人轉變。這個過程從受精卵開始,經歷蝌蚪階段,最終完成變態成為成體青蛙。這種戲劇性的的發育過程不僅在形態上引人注目,也涉及複雜的生理和生態適應。
蛙類的幼體發育通常始於水中的受精卵。卵細胞迅速分裂,形成胚胎。在適宜的溫度和濕度條件下,胚胎發育通常在幾天內完成。例如,台灣常見的盤古蟾蜍(Bufo bankorensis)的卵在水溫約25°C的條件下,大約3-4天就能孵化。在這個階段,胚胎依靠卵黃提供營養,同時發展出基本的器官系統。
孵化後,幼體進入蝌蚪階段。初孵化的蝌蚪通常十分微小,體型可能不足1厘米。它們最初仍依賴卵黃囊提供營養,但很快就會開始主動覓食。蝌蚪階段的特徵包括扁平的魚形體型、強大的尾部用於游泳、外鰓呼吸(後來發展為內鰓),以及特化的口器用於刮食藻類和其他微小有機物。
蝌蚪的發育過程可以分為幾個關鍵階段。首先是生長期,蝌蚪迅速增大體型,發展更強壯的肌肉系統。接著,後肢開始萌芽和發育,這通常是蝌蚪發育的一個重要標誌。後肢的發育速度因物種而異,有些物種的後肢發育得非常緩慢,而其他物種則相對迅速。例如,某些樹蛙的蝌蚪可能在幾週內就長出明顯的後肢。
隨著後肢的成長,蝌蚪開始進入pre-metamorphosis(前變態)階段。在這個階段,內部器官系統開始重組,為即將到來的陸地生活做準備。肺部開始發育,為未來的氣體交換做準備。同時,消化系統也開始轉變,從適應植物性飲食的長腸結構逐漸變為適應肉食的短腸結構。
接下來是變態(metamorphosis)階段,這是蛙類幼體發育中最戲劇性的的階段。前肢開始突破皮膚生長出來,尾部逐漸被吸收,頭部和口器開始重塑。這個階段的變化非常迅速,通常在幾天內就能完成。例如,台灣的莫氏樹蛙(Rhacophorus moltrechti)的蝌蚪在開始變態後,大約一週內就能完成從蝌蚪到幼蛙的轉變。
在變態過程中,蝌蚪的內部生理系統也經歷了巨大的變化。呼吸系統從鰓呼吸轉變為肺呼吸和皮膚呼吸的組合。循環系統重組,適應新的呼吸方式。同時,排泄系統也發生變化,從適應水生環境的氨排泄轉變為更適合陸地生活的尿素排泄。
值得注意的是,蛙類幼體的發育速度和持續時間因物種和環境條件而異。某些物種的蝌蚪階段可能只持續幾週,而其他物種則可能需要幾個月甚至更長時間。例如,生活在臨時水體中的物種通常有較快的發育速度,以適應水體可能乾涸的風險。相反,生活在永久性水體中的物種可能有更長的蝌蚪期,允許它們達到更大的體型後再變態。
環境因素對蛙類幼體發育有顯著的影響。溫度是一個關鍵因素,較高的溫度通常會加速發育過程。食物可用性也很重要,充足的食物可以促進更快的生長和發育。此外,捕食壓力、競爭和水質等因素也會影響發育速度和存活率。例如,在有魚類捕食者存在的水體中,某些物種的蝌蚪可能會加速變態過程,以減少被捕食的風險。
某些蛙類物種展現出幼體發育的特殊適應。例如,有些樹蛙的蝌蚪會在微小的水體中發育,如樹洞或葉片之間積累的水。這些蝌蚪通常有加速的發育過程,以適應有限的水資源。另一個極端的例子是直接發育,某些物種完全跳過了自由游泳的蝌蚪階段,直接在卵內完成所有發育過程,孵化出小青蛙。
蛙類幼體發育的研究不僅有助於我們理解這些迷人的生物的生活史,還為生態系統健康評估提供了重要工具。由於蝌蚪對環境變化非常敏感,它們常被用作生態指示物種。例如,通過監測特定水體中蝌蚪的生長速度、畸形率和存活率,科學家們可以評估水質狀況和環境污染程度。
理解蛙類幼體發育對於制定有效的保護策略至關重要。例如,知道特定物種的蝌蚪需要多長時間完成變態,可以幫助保護人員確定需要維護臨時水體的最短時間。同樣,了解不同發育階段的特定需求,可以指導棲息地管理和恢復工作,確保為蛙類提供完整的生活史所需的多樣化環境。
7.4 蛙類親代照顧
蛙類的親代照顧行為是一個令人著迷的研究領域,展現了這些兩棲動物驚人的適應能力和行為多樣性。雖然傳統觀念認為大多數蛙類在產卵後就不再關心後代,但實際上,許多物種已經演化出複雜而多樣的親代照顧策略。這些策略不僅增加了後代的存活率,也為我們理解動物行為的演化提供了寶貴的見解。
親代照顧在蛙類中的表現形式多種多樣,從最基本的選擇安全的產卵地點,到更為複雜的直接餵養和保護行為。最常見的親代照顧形式是選擇適當的產卵環境。例如,許多樹蛙物種會精心選擇樹葉或樹洞作為產卵場所。這些位置通常能提供適當的濕度和保護,減少卵被捕食或乾涸的風險。台灣的面天樹蛙(Kurixalus idiootocus)就常在樹葉背面產卵,這種行為雖然簡單,但對卵的存活至關重要。
更進一步的親代照顧表現為對卵的直接保護。某些蛙類物種的雄性或雌性會守護卵堆,防止捕食者接近或清除可能導致卵腐爛的真菌。例如,臺灣山椒魚(Hynobius formosanus)的雄性會在產卵後持續守護卵堆數週之久,直到幼體孵化。這種行為大大提高了後代的存活機會。
一些蛙類採取了更為主動的親代照顧策略,如背負卵或幼體。這種行為在負子蛙科(Atidae)的成員中尤為常見。雄性負子蛙會將受精卵串纏繞在後腿或背部,並保持卵的濕潤,直到蝌蚪準備孵化。這種策略不僅保護了卵免受捕食和乾燥,還能使親代將卵攜帶到最適宜的水體中孵化。
更為極端的親代照顧形式是體內孵化。某些蛙類物種已經演化出在體內孵化卵的能力。例如,智利袋蟾(Rhinoderma darwinii)的雄性會將受精卵吞入口中的聲囊中孵化。在這個過程中,親代不僅提供保護,還通過皮膚分泌物為發育中的幼體提供營養。這種高度專門化的親代照顧策略雖然降低了生育率,但大大提高了每個後代的存活機會。
某些樹蛙物種展示了另一種獨特的親代照顧形式:為蝌蚪提供食物。例如,熱帶美洲的毒箭蛙(Dendrobates)屬的一些物種會定期回到產卵地點,為發育中的蝌蚪提供未受精的卵作為食物。這種行為不僅確保了幼體獲得高質量的營養,還加強了親代與後代之間的聯繫。
有趣的是,蛙類的親代照顧並不總是由單一性別承擔。在不同物種中,可能是雄性、雌性或兩性共同參與照顧後代。例如,在某些樹蛙物種中,雄性負責選擇和準備產卵場所,而雌性則負責卵的護理。這種分工合作的親代照顧策略反映了複雜的演化適應。
蛙類親代照顧的程度和形式往往與其生活環境密切相關。生活在高風險環境中的物種,如乾旱地區或捕食壓力大的區域,通常會表現出更為投入的親代照顧行為。相反,在資源豐富、風險較低的環境中,親代照顧可能較為有限。這種關聯反映了親代照顧策略是如何受到環境壓力塑造的。
值得注意的是,親代照顧行為在蛙類中的演化並非單一方向。某些原本具有複雜親代照顧行為的系統可能因環境變化而退化。例如,某些原本生活在陸地上並提供直接照顧的物種,在遷移到水生環境後可能逐漸失去這些行為。這種演化的靈活性展示了蛙類如何適應不同的生態位。
蛙類親代照顧行為的研究不僅對理解這些動物的生態和演化很重要,還為保護工作提供了關鍵信息。了解特定物種的親代照顧需求可以幫助設計更有效的保護策略。例如,對於需要特定類型的產卵場所的物種,保護這些微棲息地就變得尤為重要。
此外,親代照顧行為的研究還為我們提供了洞察動物認知和學習能力的窗口。某些形式的親代照顧,如識別和餵養特定的後代,需要相當複雜的認知能力。這些發現挑戰了我們對兩棲動物智力的傳統認知。
人類活動對蛙類親代照顧行為的影響也是一個值得關注的問題。棲息地破壞、氣候變化和污染等因素可能干擾正常的親代照顧過程。例如,水質污染可能影響親代選擇產卵地點的能力,或減少可用的安全產卵場所。了解這些影響對於制定有效的保護措施至關重要。
研究蛙類的親代照顧行為不僅豐富了我們對這些迷人的生物的理解,還為更廣泛的生態學和演化生物學問題提供了見解。通過研究蛙類如何平衡投資和回報,我們可以更好地理解親代照顧策略在自然界中的演化動態。這些知識不僅對科學研究有價值,也對生物多樣性保護和生態系統管理具有實際意義。
7.5 蛙類的獨特繁殖適應
蛙類在長期的演化過程中發展出了許多獨特而迷人的的繁殖適應策略。這些適應不僅展現了自然選擇的創造力,也反映了蛙類對各種環境挑戰的應對能力。從特殊的產卵方式到驚人的胚胎發育過程,蛙類的繁殖適應涵蓋了一系列令人驚嘆的創新。
其中最引人注目的繁殖適應之一是某些物種的泡沫巢。例如,生活在熱帶和亞熱帶地區的泡沫巢蛙科(Rhacophoridae)成員,會在水面上或樹枝上製造一個由泡沫組成的巢穴。雌蛙在產卵過程中會分泌一種特殊的蛋白質,當這種蛋白質與空氣接觸並被雄蛙的後腿攪拌時,就會形成泡沫。這個泡沫巢不僅能保護卵免受捕食和乾燥,還能提供一個穩定的溫度和濕度環境。更令人驚奇的是,當蝌蚪準備孵化時,泡沫會自動溶解,讓幼體安全地進入水中。這種適應特別適合在乾旱或半乾旱地區繁殖的蛙類,因為它能最大限度地減少水分流失。
另一種獨特的繁殖適應是背部育幼袋。澳洲的袋蟾蜍(Assa darlingtoni)就展示了這種令人驚嘆的適應。在這個物種中,雄性背部的皮膚會形成特殊的口袋狀結構。雌性將卵產在地面上後,雄性會用後腿將卵掃入這些背部口袋中。卵在口袋中發育,直接變態為小蛙後才離開雄性的背部。這種適應不僅提供了極高程度的親代保護,還允許蛙類在缺乏永久性水體的環境中成功繁衍。
直接發育是另一種顯著的繁殖適應。某些生活在潮濕熱帶環境中的蛙類完全跳過了自由游泳的蝌蚪階段。例如,某些小型地棲蛙類的胚胎在卵內完成全部發育過程,直接孵化為微小的青蛙。這種適應消除了對水體的依賴,使這些蛙類能夠在森林地面等缺乏開放水域的環境中繁衍。直接發育的一個極端例子是所羅門群島的所羅門島蛙(Ceratobatrachus guentheri),其卵中的胚胎發育出特殊的呼吸器官,能夠直接從卵壳吸收氧氣。
某些蛙類發展出了體內受精的能力,這在兩棲動物中相當罕見。例如,非洲的有尾蛙(Ascaphus truei)雄性具有一個類似尾巴的交配器官,用於內部受精。這種適應允許蛙類在湍急的溪流中成功繁殖,因為外部受精在這種環境下很難實現。
胃孵化是另一種極其獨特的繁殖適應。已經滅絕的澳洲胃孵蛙(Rheobatrachus silus)就曾展示了這種驚人的能力。雌蛙會將受精卵吞入胃中,並停止分泌胃酸。卵在胃中發育為蝌蚪,最後變態為小蛙從母體口中「出生」。這種適應提供了最高程度的親代保護,但也要求母體做出重大的生理調整。
某些樹蛙物種發展出了「空中繁殖」的能力。例如,有些樹蛙會在高處的樹葉上產卵,卵會發育成蝌蚪,然後直接掉入下方的水體中。這種策略既避免了水中的許多捕食者,又確保了蝌蚪能夠進入適合的水生環境。
「反向」性別角色也是一種有趣的繁殖適應。在大多數蛙類中,雄性通過鳴叫吸引雌性。然而,在某些物種中,如非洲的弓蛙(Hyperolius argus),這個角色被顛倒了。雌性會主動鳴叫吸引雄性,並且在求偶過程中表現得更為積極。這種適應可能與特定的環境壓力或繁殖資源的分布有關。
某些蛙類還發展出了特殊的卵結構作為繁殖適應。例如,玻璃蛙科(Centrolenidae)的成員產下透明的卵,這些卵通常附著在懸掛在水面上的葉子底部。透明的卵不僅能夠偽裝,還允許陽光穿透,可能加速胚胎發育。
蛙類的繁殖適應還包括對極端環境的適應。例如,生活在沙漠邊緣的某些蟾蜍物種能夠在極短的時間內完成繁殖過程。它們可能整年蟄伏在地下,只在短暫的雨季出來繁殖。這些蛙類的卵和蝌蚪發育速度極快,以適應短暫的水體環境。
值得注意的是,這些獨特的繁殖適應往往伴隨著生理和行為上的特殊變化。例如,某些提供高度親代照顧的物種可能會降低每次繁殖的後代數量,但增加每個後代的存活機會。這反映了繁殖策略中的權衡取捨。
研究蛙類的這些獨特繁殖適應不僅能增進我們對生物多樣性的理解,還為生物技術和醫學研究提供了靈感。例如,泡沫巢蛙的泡沫蛋白質結構研究可能為開發新型生物材料提供線索。同時,這些研究對於制定有效的蛙類保護策略也至關重要,因為不同的繁殖適應可能需要不同的保護方法。

8. 蛙類覓食生態學
蛙類的覓食生態學是一個引人入勝的研究領域,揭示了這些兩棲動物如何適應不同環境並維持生存。蛙類的食性組成相當多樣,從小型昆蟲到其他小型脊椎動物都可能成為牠們的獵物。不同種類的蛙採用各式各樣的覓食策略,有些會靜候獵物接近,有些則主動追捕。蛙類捕食機制中最為人熟知的是其獨特的黏性舌頭,能夠迅速彈出捕獲獵物。
某些蛙類還發展出特殊的飲食適應,例如有些樹蛙的嘴巴結構特化,適合捕食飛行中的昆蟲。水生蛙類則可能以水生昆蟲或小魚為食。值得注意的是,蛙類在生態系統中扮演著重要的角色,特別是在害蟲控制方面。在農業地區,蛙類能夠有效地控制蚊子、蒼蠅等有害昆蟲的數量,從而減少農作物的損失和疾病的傳播。
然而,蛙類的覓食行為也面臨著人為活動帶來的挑戰。農藥的使用不僅直接影響蛙類的健康,還會減少牠們的食物來源。此外,棲息地的改變也可能導致食物鏈的變動,進而影響蛙類的覓食模式。因此,了解蛙類的覓食生態學不僅對於保護這些物種至關重要,也有助於維護整個生態系統的平衡。
8.1 蛙類食性組成
蛙類的食性組成是一個複雜而多樣的主題,反映了這些兩棲動物在適應不同生態環境中的進化過程。蛙類作為生態系統中的重要成員,其食性組成不僅影響自身的生存和繁衍,還對整個生態系統的平衡起著關鍵作用。
一般而言,大多數蛙類是肉食性動物,主要以各種小型無脊椎動物為食。昆蟲是蛙類食譜中最常見的組成部分,包括蒼蠅、蚊子、甲蟲、蝴蝶和蛾類等。這些昆蟲不僅為蛙類提供了豐富的蛋白質來源,還包含了必要的礦物質和維生素。除了昆蟲外,蜘蛛、蠕蟲、蝸牛和其他軟體動物也經常出現在蛙類的食譜中。
然而,蛙類的食性組成並非僅限於無脊椎動物。一些較大型的蛙類,如非洲牛蛙(Pyxicephalus adspersus)和角蛙(Ceratophrys ornata),甚至能夠捕食小型脊椎動物,包括其他蛙類、蜥蜴、小鳥和齧齒類動物。這種食性的多樣性展示了蛙類在食物鏈中的不同位置,從初級消費者到高級掠食者都有蛙類的身影。
值得注意的是,蛙類的食性組成會隨著其生命週期的不同階段而變化。蝌蚪階段的蛙類主要以藻類、水生植物碎屑和微小的水生生物為食。隨著變態的進行,蝌蚪逐漸轉向肉食性飲食。這種飲食習慣的轉變不僅反映了蛙類形態結構的變化,也體現了其在生態系統中角色的轉變。
蛙類的食性組成還會因季節變化而有所不同。在食物豐富的季節,蛙類可能會選擇更為多樣的食物來源。相反,在食物稀缺的時期,它們可能會降低對食物種類的選擇性,以確保能夠攝取足夠的營養。例如,研究發現某些樹蛙在雨季時主要捕食飛行昆蟲,而在乾季則更多地依賴地面爬行的昆蟲。
地理位置和棲息環境也是影響蛙類食性組成的重要因素。生活在熱帶雨林中的蛙類可能有更多樣化的食物選擇,而生活在乾旱或寒冷地區的蛙類則可能面臨更有限的食物來源。例如,生活在南美洲亞馬遜雨林的箭毒蛙(Dendrobatidae)主要以螞蟻和白蟻為食,這些昆蟲不僅提供了營養,還是箭毒蛙毒素的來源。
某些蛙類還表現出特殊的飲食偏好或適應。例如,非洲爪蟾(Xenopus laevis)具有高度特化的濾食系統,能夠在水中濾食浮游生物和有機碎屑。這種獨特的飲食方式使得非洲爪蟾能夠在各種水體中生存,甚至包括污染較為嚴重的水域。
人類活動對蛙類的食性組成也產生了顯著影響。城市化和農業活動改變了許多蛙類的原有棲息地,導致其食物來源發生變化。一些適應能力較強的蛙類,如綠樹蛙(Hyla cinerea),已經學會利用人造光源吸引的昆蟲作為食物來源。然而,這種行為可能使蛙類更容易暴露於捕食者的威脅之下。
農藥的使用是另一個影響蛙類食性組成的重要人為因素。殺蟲劑不僅直接減少了蛙類的食物來源,還可能通過食物鏈的生物累積對蛙類造成毒害。這種情況不僅威脅蛙類的生存,還可能導致生態系統的失衡。
研究蛙類的食性組成對於理解和保護這些兩棲動物至關重要。通過分析蛙類的糞便、胃內容物或利用穩定同位素技術,科學家們能夠更精確地了解不同蛙類的食性組成。這些研究不僅有助於我們理解蛙類在生態系統中的角色,還為制定有效的保護策略提供了重要依據。
此外,蛙類的食性組成研究還為生物防治提供了新的思路。某些蛙類對特定害蟲的偏好使得它們成為潛在的生物防治工具。例如,在一些水稻種植區,利用蛙類控制害蟲的數量已經取得了一定的成效。
了解蛙類的食性組成不僅對生態學研究具有重要意義,還為我們提供了觀察自然界複雜相互作用的窗口。蛙類作為連接水陸生態系統的重要紐帶,其食性組成的研究揭示了生態系統中能量流動和物質循環的奧秘。隨著研究方法的不斷進步和對蛙類生態學認識的深入,我們相信未來會有更多關於蛙類食性組成的新發現,這將為生態保護和環境管理提供更多的科學依據。
8.2 蛙類覓食策略
蛙類的覓食策略是一個引人入勝的話題,反映了這些兩棲動物在長期進化過程中為適應不同環境而發展出的多樣化生存技能。蛙類的覓食策略不僅涉及獵捕行為,還包括獵物選擇、時間分配和能量利用等多個方面,展現了蛙類在生態系統中的適應性和重要性。
最常見的蛙類覓食策略是「守株待兔」式的坐等捕食。許多蛙類會選擇一個有利位置,靜靜地等待獵物接近。這種策略特別適合那些體色與環境相近的蛙類,如樹蛙和某些地棲蛙類。牠們通常具有出色的保護色,能夠完美地融入周圍環境。例如,綠色的樹蛙常常棲息在樹葉或草叢中,而棕色的地棲蛙則喜歡藏身於落葉堆中。這種靜待策略不僅能夠節省能量,還能有效地避開天敵的注意。
然而,並非所有蛙類都採用被動等待的策略。一些較為活躍的蛙類,如赤蛙科的某些成員,會主動搜尋獵物。這些蛙類通常具有較強的運動能力,能夠在地面或水中快速移動。牠們會在一定範圍內巡視,尋找可能的獵物。這種主動搜尋的策略雖然消耗更多能量,但能夠增加覓食成功的機會,特別是在食物資源相對稀少的環境中。
一些特化的蛙類還發展出了獨特的覓食策略。例如,非洲爪蟾(Xenopus laevis)採用了一種名為「吸入式捕食」的方法。這種水生蛙類能夠通過快速張開口腔,產生負壓將水和獵物一起吸入口中。這種策略特別適合在渾濁的水中捕捉小型獵物,如水蚤和其他浮游生物。
某些樹蛙則採用了「伏擊式」的捕食策略。牠們會選擇在樹枝或樹葉的下方倒掛著,等待獵物從下方經過。當獵物接近時,蛙類會迅速跳下抓住獵物。這種策略不僅利用了重力優勢,還能夠有效地避開某些天敵的視線。
值得注意的是,蛙類的覓食策略常常與其形態特徵密切相關。例如,具有大眼睛和寬大口部的蛙類通常更擅長捕捉飛行中的昆蟲。相比之下,那些具有較小眼睛和窄長身體的蛙類可能更適合在地面或水中捕捉爬行的獵物。
時間也是蛙類覓食策略的一個重要因素。大多數蛙類是夜行性動物,在黃昏和夜間活動。這種行為不僅可以避開日間的高溫和乾燥環境,還能夠降低被視覺型捕食者發現的風險。然而,也有一些蛙類,如箭毒蛙(Dendrobatidae),是日行性的。這些鮮艷的蛙類通常具有毒性,不需要依賴隱蔽來避免被捕食。
蛙類的覓食策略還會隨季節變化而調整。在食物豐富的季節,蛙類可能會採取更為積極的覓食策略,增加活動範圍和頻率。而在食物稀少的時期,牠們可能會降低活動頻率,更多地依賴「守株待兔」的策略以節省能量。
一些蛙類還表現出複雜的學習行為和記憶能力,這些能力在其覓食策略中發揮重要作用。研究發現,某些蛙類能夠記住高產量的覓食地點,並在未來的覓食活動中優先選擇這些位置。這種行為不僅提高了覓食效率,還展示了蛙類認知能力的複雜性。
人類活動對蛙類的覓食策略也產生了顯著影響。城市化帶來的光污染改變了某些蛙類的活動模式。一些原本夜行的蛙類開始利用街燈吸引的昆蟲作為食物來源,這種行為雖然增加了食物獲取的機會,但同時也增加了被捕食的風險。
氣候變化也迫使蛙類調整其覓食策略。全球變暖導致某些地區的昆蟲活動高峰提前,蛙類必須相應地調整其活動時間以適應這種變化。此外,極端天氣事件的增加,如乾旱或暴雨,也要求蛙類具備更靈活的覓食策略以應對不穩定的環境。
研究蛙類的覓食策略不僅有助於我們理解這些生物的生態學特性,還為保護瀕危蛙類提供了重要依據。通過了解不同蛙類的覓食需求和行為模式,我們可以更好地設計和管理保護區,確保這些區域能夠提供蛙類所需的多樣化食物資源和適宜的覓食環境。
此外,蛙類的覓食策略研究還為生物防治提供了新的思路。某些蛙類對特定害蟲的偏好使得它們成為潛在的生物防治工具。例如,在一些水稻種植區,利用蛙類控制害蟲的數量已經取得了一定的成效。通過深入了解蛙類的覓食行為和偏好,我們可以更有效地利用這些天然的害蟲控制者。
蛙類覓食策略的多樣性和適應性不僅體現了自然選擇的力量,還展示了生態系統中複雜的相互作用。作為連接水陸生態系統的重要紐帶,蛙類的覓食行為對於維持生態平衡起著關鍵作用。隨著研究方法的不斷進步和對蛙類生態學認識的深入,我們相信未來會有更多關於蛙類覓食策略的新發現,這將為生態保護和環境管理提供更多的科學依據。
8.3 蛙類捕食機制
蛙類的捕食機制是一個令人著迷的生物學主題,展現了這些兩棲動物在進化過程中發展出的精妙適應。蛙類捕食機制的核心在於其獨特的解剖結構和行為模式,這些特徵使得蛙類能夠高效地捕獲各種獵物。
最為人熟知的蛙類捕食機制是利用其特化的舌頭。蛙類的舌頭結構獨特,通常附著在口腔前部,而非後部。這種構造使得舌頭能夠像彈簧一樣快速彈出。當蛙類發現獵物時,它們能在極短的時間內(通常不到0.07秒)將舌頭彈出,準確地黏住獵物並將其拉回口中。這個過程快到人眼幾乎難以捕捉。
蛙類舌頭的黏性是其捕食成功的關鍵。研究發現,蛙類舌頭表面覆蓋著一層特殊的黏液,這種黏液具有非牛頓流體的特性。當舌頭接觸獵物時,黏液會瞬間變得更加黏稠,確保獵物被牢牢黏住。同時,舌頭表面的微小凹凸結構增加了與獵物接觸的表面積,進一步提高了捕獲效率。
除了舌頭,蛙類的口腔結構也在捕食過程中扮演重要角色。許多蛙類具有可以快速下降的上顎,這種機制被稱為「口腔泵」(buccal pumping)。當舌頭將獵物帶回口腔時,上顎迅速下降,產生負壓,協助將獵物吸入口中。這個過程通常伴隨著眼球的下沉,因為眼球與口腔相連,眼球的下壓有助於增加口腔容積,進一步增強吸力。
某些水生蛙類,如非洲爪蟾(Xenopus laevis),則發展出了不同的捕食機制。這些蛙類通過快速擴張口腔和喉部,在水中產生強大的吸力,將水和獵物一起吸入口中。這種「抽水泵」式的捕食方法特別適合在渾濁的水體中捕捉小型獵物。
蛙類的視覺系統在捕食過程中也起著關鍵作用。大多數蛙類都具有突出的大眼睛,提供了寬廣的視野。蛙類的眼睛能夠快速適應光線變化,這使得它們在不同光照條件下都能有效捕食。有趣的是,研究發現蛙類主要對移動的物體產生反應,這可能是為了在複雜的自然環境中更好地識別潛在獵物。
一些蛙類還具有特殊的捕食適應。例如,某些樹蛙的腳趾末端有吸盤狀結構,這不僅有助於它們在樹枝上攀爬,還能在捕食時提供額外的支撐,使得它們能夠在各種角度和位置精確地捕獲獵物。
大型的蛙類,如非洲牛蛙(Pyxicephalus adspersus)和角蛙(Ceratophrys ornata),則採用一種更為直接的捕食方法。這些蛙類通常具有巨大的口部和強壯的下顎肌肉,能夠直接用嘴咬住並吞下較大的獵物,包括其他小型脊椎動物。
蛙類的捕食機制還與其身體其他部位的協調動作密切相關。例如,在捕食過程中,蛙類的前肢常常用來協助將大型獵物塞入口中或調整獵物的位置。有些蛙類甚至會用前肢將過大的獵物撕成小塊,以便吞咽。
值得注意的是,蛙類的捕食機制會隨著其生命階段的變化而改變。蝌蚪階段的蛙類主要依靠過濾式攝食,利用口部特化的結構過濾水中的微小有機物。隨著變態的進行,蛙類逐漸發展出成體的捕食機制。這種轉變不僅涉及形態結構的改變,還包括神經系統和行為模式的調整。
環境因素也會影響蛙類的捕食機制。例如,在光線不足的環境中,某些蛙類可能更多地依賴聽覺和觸覺來定位獵物。一些生活在洞穴或地下環境的蛙類甚至進化出了特殊的感覺器官,以彌補視覺的不足。
人類活動對蛙類的捕食機制也產生了影響。環境污染,特別是水體污染,可能會影響水生蛙類的「抽水泵」式捕食效率。農藥的使用不僅直接威脅蛙類的健康,還可能改變獵物的行為模式,間接影響蛙類的捕食成功率。
研究蛙類的捕食機制不僅有助於我們理解這些生物的生態學特性,還為生物工程和機器人技術提供了靈感。例如,蛙類舌頭的快速運動和黏附特性已經被應用於開發新型抓取裝置。此外,蛙類的視覺系統和運動協調能力也為計算機視覺和機器人運動控制提供了有價值的參考。
深入了解蛙類的捕食機制對於保護這些物種也具有重要意義。通過了解不同蛙類的捕食需求和行為模式,我們可以更好地設計和管理保護區,確保這些區域能夠提供蛙類所需的多樣化食物資源和適宜的捕食環境。這對於維護生態系統的平衡和生物多樣性保護都具有深遠的意義。
8.4 不同蛙種的飲食適應
不同蛙種的飲食適應是蛙類生態學中一個引人入勝的主題,反映了這些兩棲動物在長期進化過程中為適應各種生態環境而發展出的多樣化生存策略。蛙類的飲食適應不僅涉及食物選擇,還包括形態結構、生理機能和行為模式等多個方面的特化,展現了自然選擇的神奇力量。
樹蛙科(Hylidae)的成員是飲食適應的典型代表。這些蛙類通常棲息在樹上或灌木叢中,其飲食適應主要體現在捕捉飛行昆蟲的能力上。樹蛙的眼睛通常較大,視力敏銳,能夠在複雜的樹冠環境中準確定位飛行中的獵物。它們的舌頭特別黏滑,能夠快速彈出並精確捕獲空中的昆蟲。此外,樹蛙的腳趾末端有吸盤狀結構,不僅有助於攀爬,還能在捕食時提供穩定支撐,使得它們能夠在各種角度和位置精確地捕獲獵物。
相比之下,生活在地面或水邊的赤蛙科(Ranidae)成員則展現出不同的飲食適應。這些蛙類通常以地面爬行的昆蟲、蠕蟲和其他小型無脊椎動物為食。赤蛙的後腿肌肉發達,使它們能夠快速跳躍捕捉獵物。它們的舌頭結構也適合捕捉地面獵物,通常較為寬大,能夠覆蓋較大的區域。某些水邊生活的赤蛙還能捕食小魚,它們的消化系統適應了處理含有魚刺的食物。
蟾蜍科(Bufonidae)的成員則展現出另一種獨特的飲食適應。蟾蜍通常以小型節肢動物為食,特別是螞蟻和甲蟲。它們的舌頭相對較短但肌肉發達,適合捕捉地面上的小型獵物。蟾蜍的皮膚分泌物具有毒性,這不僅是防禦機制,還能夠在某種程度上影響它們的飲食選擇,使得一些通常會捕食蛙類的掠食者避而遠之。
箭毒蛙科(Dendrobatidae)的飲食適應尤為特別。這些色彩鮮艷的小型蛙類主要以螞蟻和白蟻為食,它們的毒性實際上來源於所食用的昆蟲。箭毒蛙的消化系統能夠將這些昆蟲體內的毒素積累並改造,形成自身的防禦武器。這種獨特的飲食適應不僅影響了箭毒蛙的生理特性,還塑造了它們的行為模式和生態位。
水生蛙類,如非洲爪蟾(Xenopus laevis),則發展出了完全不同的飲食適應。這些蛙類通過快速擴張口腔和喉部,在水中產生強大的吸力,將水和獵物一起吸入口中。它們的指尖有利爪狀結構,有助於在水中抓取和操縱獵物。非洲爪蟾的消化系統適應了處理水生小動物,如水蚤、蚊蟲幼蟲等。
某些大型蛙類,如非洲牛蛙(Pyxicephalus adspersus)和角蛙(Ceratophrys ornata),展現出捕食大型獵物的適應。這些蛙類具有巨大的口部和強壯的下顎肌肉,能夠捕食其他蛙類、蜥蜴、小鳥甚至小型哺乳動物。它們的消化系統能夠處理包含骨骼在內的大型獵物,這種適應使得這些蛙類在食物鏈中處於較高的位置。
狹口蛙科(Microhylidae)的成員則展現出高度特化的飲食適應。這些蛙類通常體型較小,口部狹窄,特別適合捕食螞蟻和白蟻。它們的舌頭結構獨特,能夠深入蟻穴捕獲獵物。一些狹口蛙甚至發展出了共生關係,生活在白蟻或螞蟻的巢穴中,以這些社會性昆蟲為食。
蛙類的飲食適應還體現在其消化系統的特化上。例如,以硬殼甲蟲為主要食物的蛙類通常具有較厚的胃壁和更強的胃酸分泌能力,以便消化堅硬的甲殼。而以軟體動物為食的蛙類則可能具有特殊的酶系統,能夠有效分解這類獵物的黏液和軟組織。
值得注意的是,蛙類的飲食適應並非固定不變的。許多蛙類展現出根據環境變化調整飲食習慣的能力。例如,在食物稀缺的季節,原本以昆蟲為主食的蛙類可能會轉而捕食小型脊椎動物或其他通常不在其食譜中的生物。這種飲食靈活性是蛙類適應多變環境的重要策略。
蛙類的飲食適應還與其生活環境密切相關。生活在乾旱地區的蛙類可能會發展出儲存水分的能力,並傾向於選擇含水量高的獵物。而生活在高海拔或寒冷地區的蛙類可能會發展出更高效的能量利用機制,選擇能量密度較高的食物。
人類活動對蛙類的飲食適應也產生了顯著影響。城市化導致某些蛙類不得不適應新的食物來源,如利用路燈吸引的昆蟲。農業活動的擴張也改變了許多蛙類的飲食模式,有些蛙類開始以農作物害蟲為食,在某種程度上成為了農業生態系統中的天然害蟲控制者。
研究不同蛙種的飲食適應不僅有助於我們理解生態系統的複雜性,還為保護瀕危蛙類提供了重要依據。通過了解特定蛙類的飲食需求,我們可以更好地設計和管理保護區,確保這些區域能夠提供適當的食物資源。此外,這些研究還為生物防治和生態工程提供了新的思路,例如利用特定蛙類控制農業害蟲或管理城市生態系統。
8.5 蛙類在害蟲控制中的角色
蛙類在生態系統中扮演著重要的角色,尤其是在害蟲控制方面,它們的貢獻常常被低估。作為自然界中的捕食者,蛙類在維持生態平衡和控制有害生物數量方面發揮著關鍵作用。這種生態功能不僅對自然生態系統至關重要,還對農業生產和人類健康有著深遠的影響。
在農業生態系統中,蛙類是許多農作物害蟲的天敵。它們的食譜中包括多種農業有害生物,如蚜蟲、蝗蟲、蛾類幼蟲等。例如,在水稻田中,青蛙(Rana limnocharis)被認為是一種有效的生物防治工具。這種蛙類能夠捕食多種水稻害蟲,包括稻飛虱、稻蝗和某些蛾類的幼蟲。研究表明,在水稻田中保持適當的青蛙數量可以顯著減少農藥的使用,同時提高作物產量。
除了直接捕食害蟲,蛙類的存在還能夠影響害蟲的行為。害蟲為了避免被捕食,可能會減少活動或改變覓食模式,從而間接降低對作物的危害。這種非消耗性效應(non-consumptive effect)雖然難以量化,但在整體的害蟲控制中扮演著重要角色。
蛙類在控制蚊子數量方面也有顯著貢獻。許多蛙類,特別是那些生活在水邊或濕地的種類,會捕食蚊子的幼蟲和成蟲。例如,綠蛙(Rana clamitans)和美洲牛蛙(Lithobates catesbeianus)都是蚊子的有效天敵。通過控制蚊子數量,蛙類間接地幫助減少了一些由蚊子傳播的疾病,如瘧疾、登革熱和寨卡病毒等。
在城市環境中,蛙類同樣在害蟲控制方面發揮作用。城市公園、花園和其他綠地中的蛙類能夠幫助控制各種昆蟲的數量,包括一些被視為城市害蟲的物種。例如,樹蛙(Hyla spp.)常常在城市環境中發現,它們能夠有效地捕食蚊子、蛾類和其他飛行昆蟲。
然而,蛙類在害蟲控制中的作用並非僅限於直接捕食。它們在食物鏈中的位置使得它們成為生態系統健康的重要指標。蛙類數量的變化可以反映出整個生態系統的平衡狀況,包括害蟲和其他生物的數量變化。因此,監測蛙類種群可以為農業和生態管理提供寶貴的信息。
蛙類的害蟲控制能力與其獨特的生理和行為特徵密切相關。例如,許多蛙類具有寬大的口部和黏性強的舌頭,能夠捕獲各種大小的昆蟲。它們的夜行性習慣使得它們能夠捕食許多在夜間活動的害蟲,這些害蟲可能逃過了日間活動的鳥類的捕食。
值得注意的是,不同種類的蛙類在害蟲控制中可能扮演不同的角色。例如,較大型的蛙類如牛蛙可能更擅長捕食大型昆蟲,而小型的樹蛙則可能更適合控制飛行昆蟲。這種多樣性使得蛙類群落能夠更全面地控制各種害蟲。
蛙類在害蟲控制中的作用還體現在它們的生活史特徵上。許多蛙類在幼體(蝌蚪)階段主要以藻類和有機碎屑為食,而成體則轉為捕食昆蟲。這種生活史策略使得蛙類能夠在不同的生態位上發揮作用,既能夠通過蝌蚪階段控制水體中的藻類生長,又能在成體階段控制陸地和水生昆蟲的數量。
在有機農業和生態農業中,蛙類的害蟲控制作用受到越來越多的重視。一些農民開始有意識地在農田周圍創造適合蛙類生存的環境,如保留小型水體或濕地,以吸引和維持蛙類種群。這種做法不僅有助於害蟲控制,還能提高農田生態系統的整體生物多樣性。
然而,蛙類在害蟲控制中的作用也面臨著挑戰。農藥的廣泛使用是一個主要問題。許多農藥不僅殺死害蟲,也會對蛙類造成傷害,無論是直接毒性還是通過食物鏈的生物累積。這種情況不僅降低了蛙類的數量,也削弱了它們在害蟲控制中的作用。
氣候變化也對蛙類的害蟲控制功能產生影響。溫度和降水模式的改變可能影響蛙類的分布和活動模式,進而影響它們對害蟲的控制能力。例如,乾旱可能導致蛙類種群減少,從而減弱它們在害蟲控制中的作用。
棲息地破壞是另一個威脅蛙類害蟲控制功能的因素。隨著城市化和農業擴張,許多蛙類的自然棲息地被破声或片段化。這不僅直接影響蛙類的生存,還可能改變它們的覓食行為和效率。
儘管面臨這些挑戰,但蛙類在害蟲控制中的重要性正得到越來越多的認識。一些地區開始實施保護措施,如建立生態廊道、恢復濕地等,以維持和增強蛙類種群。同時,一些農業實踐也開始注重與蛙類友好的方法,如減少農藥使用、保留田邊水體等。
研究蛙類在害蟲控制中的作用不僅有助於我們更好地理解和管理農業生態系統,還為生態友好型的害蟲管理策略提供了新的思路。通過合理利用蛙類的這一生態功能,我們有望在未來實現更加可持續和環境友好的農業生產模式。

9. 蛙類的種間互動
蛙類的種間互動是生態系統中複雜而有趣的一環,展現了生物多樣性的重要性。這些互動包括捕食關係、競爭、共生以及蛙類作為生物指標的角色。在食物網中,蛙類既是捕食者也是獵物,它們捕食昆蟲和小型無脊椎動物,同時也被鳥類、蛇類和哺乳動物獵食。這種雙重角色使蛙類成為生態系統能量流動的重要媒介。
在資源有限的環境中,不同種類的蛙類之間存在競爭關係,特別是在繁殖地和食物資源方面。例如,在同一水體中,不同蛙種可能為了最佳的產卵位置而競爭。一些蛙類還與其他生物形成了互利共生關係,如某些樹蛙與植物之間的關係,蛙類為植物提供養分,同時獲得棲息場所。
蛙類作為環境變化的敏感指標,其種群動態可以反映生態系統的健康狀況。例如,某些蛙類對水質變化特別敏感,其數量變化可以早期預警水體污染。此外,一些蛙類還在授粉和種子傳播中扮演著意想不到的角色,尤其是在熱帶雨林生態系統中。這些多樣的種間互動凸顯了蛙類在維持生態平衡中的重要性,也為我們理解和保護生物多樣性提供了重要視角。
9.1 蛙類捕食者-獵物動態
蛙類在生態系統中扮演著獨特的雙重角色,既是捕食者也是獵物,這使得蛙類的捕食者-獵物動態成為生態學研究中一個引人入勝的主題。這種複雜的關係不僅影響蛙類自身的種群動態,還對整個生態系統的平衡產生深遠影響。
作為捕食者,蛙類主要以各種小型無脊椎動物為食。大多數蛙類偏好捕食昆蟲,如蚊子、蒼蠅、甲蟲和蝴蝶等。這種捕食行為對於控制昆蟲種群數量起著重要作用。例如,在水稻田生態系統中,青蛙(Rana limnocharis)被視為重要的生物防治工具,它們能夠有效控制如稻飛虱和稻蝗等農業害蟲的數量。蛙類的捕食行為不僅直接減少害蟲數量,還能通過改變害蟲的行為模式間接影響其種群動態。
然而,蛙類自身也是眾多捕食者的目標。鳥類、蛇類、魚類和某些哺乳動物都將蛙類視為食物來源。例如,蒼鷺(Ardea cinerea)是許多水生和半水生蛙類的主要天敵。這種捕食壓力對蛙類的行為和種群動態產生顯著影響。為了應對這種壓力,許多蛙類進化出了各種防禦機制,如保護色、毒性皮膚分泌物和警戒行為等。
蛙類的捕食者-獵物動態還與其生活史階段密切相關。蝌蚪階段的蛙類主要以藻類和有機碎屑為食,同時也面臨著來自水生昆蟲、魚類和其他蝌蚪的捕食威脅。隨著變態過程的進行,蛙類的食性和面臨的捕食壓力都會發生顯著變化。這種生活史策略的轉變使得蛙類在不同的生態位上都扮演著重要角色,從而影響整個生態系統的能量流動和物質循環。
在某些生態系統中,蛙類可能成為關鍵種(keystone 物種),其種群動態的變化可能引發一系列連鎖反應。例如,在某些池塘生態系統中,大型蛙類(如牛蛙)的引入可能導致原有的小型蛙類種群下降,進而影響整個食物網的結構。這種現象突顯了蛙類在維持生態平衡中的重要性。
蛙類的捕食者-獵物動態還受到環境因素的顯著影響。季節變化、氣候條件和棲息地特徵都可能改變蛙類的捕食行為和被捕食風險。例如,在乾旱季節,水源的減少可能導致蛙類和其捕食者集中在有限的水體中,從而加劇捕食壓力。相反,在雨季,增加的水體和植被覆蓋可能為蛙類提供更多的庇護所和捕食機會。
人類活動也對蛙類的捕食者-獵物動態產生深遠影響。棲息地破壞和片段化可能改變蛙類和其捕食者的空間分布,從而影響它們之間的相互作用。農藥的使用不僅直接危害蛙類,還可能通過減少昆蟲數量間接影響蛙類的食物來源。氣候變化則可能改變蛙類和其捕食者的活動模式和分布範圍,進而影響整個食物網的結構。
在研究蛙類的捕食者-獵物動態時,科學家們發現了一些有趣的現象。例如,某些蛙類表現出對捕食者的學習能力。研究表明,經歷過捕食者攻擊的蛙類個體在未來面對類似威脅時表現出更強的警戒行為。這種學習能力不僅影響個體行為,還可能通過社會學習影響整個種群的行為模式。
另一個有趣的現象是蛙類和其捕食者之間的軍備競賽(arms race)。隨著蛙類進化出更有效的防禦機制,其捕食者也相應地進化出更強的捕食能力。例如,某些蛇類已經進化出對蛙類毒素的抗性,能夠捕食原本有毒的蛙類。這種持續的進化過程塑造了現今我們所觀察到的複雜捕食者-獵物關係。
在某些情況下,蛙類還可能表現出食腐行為,這為它們的捕食者-獵物動態增添了新的維度。例如,在食物稀缺的環境中,某些蛙類被觀察到會攝食已死亡的動物。這種行為不僅拓展了蛙類的食物來源,還可能影響生態系統中的營養循環。
蛙類的捕食者-獵物動態研究對於保護生物學和生態系統管理具有重要意義。通過了解這些複雜的相互作用,我們可以更好地預測環境變化對蛙類種群和整個生態系統的影響。例如,在設計保護區時,考慮蛙類和其捕食者的需求可以幫助創造更穩定和多樣化的生態系統。
此外,蛙類捕食者-獵物動態的研究還為生物防治提供了新的思路。通過合理利用蛙類的捕食行為,我們可能開發出更環保、更可持續的害蟲管理策略。同時,了解蛙類面臨的捕食壓力也有助於我們制定更有效的蛙類保護措施。
9.1.1 台灣最常見吃蛙類的鳥類?
台灣作為一個生物多樣性豐富的島嶼,擁有多種以蛙類為食的鳥類。這些鳥類在生態系統中扮演著重要的角色,不僅影響蛙類的種群動態,也反映了台灣獨特的生態環境特徵。
其中,最常見的吃蛙類的鳥類首推鷺科(Ardeidae)的成員。夜鷺(Nycticorax nycticorax)是台灣常見的夜行性鳥類,經常在黃昏或夜間在水邊活動,以蛙類、魚類和水生昆蟲為食。牠們的捕食行為對於控制蛙類種群數量起著重要作用。夜鷺的存在不僅影響蛙類的數量,還可能改變蛙類的行為模式,如使蛙類減少鳴叫或改變活動時間以避免被捕食。
小白鷺(Egretta garzetta)是另一種常見的捕食蛙類的鳥類。這種優雅的白色鳥類經常出現在台灣的濕地、水田和河岸地帶。小白鷺採用站立等待的捕食策略,耐心地在淺水區等待獵物出現。當發現蛙類時,牠們會迅速出擊,用尖銳的喙捕獲獵物。小白鷺的存在對維持濕地生態系統的平衡具有重要意義。
蒼鷺(Ardea cinerea)是體型較大的鷺科鳥類,同樣以蛙類為主要食物來源之一。這種鳥類具有出色的耐心和精準的捕食技巧,能夠長時間靜立在水中等待獵物。蒼鷺的長腿和長頸使牠們能夠在較深的水域中捕食,這讓牠們能夠捕獲一些較大型的蛙類,如石蛙或虎皮蛙。
除了鷺科鳥類,台灣的鷸科(Scolopacidae)鳥類中也有一些以蛙類為食的種類。青足鷸(Tringa nebularia)是一種常見的水邊鳥類,牠們在遷徙季節會大量出現在台灣的濕地和海岸地區。雖然青足鷸主要以小型魚類和水生昆蟲為食,但牠們也會捕食小型的蛙類,特別是在潮間帶或淺水區域。
台灣藍鵲(Urocissa caerulea)是台灣特有種,雖然主要以昆蟲和果實為食,但也會opportunistical捕食小型脊椎動物,包括蛙類。這種美麗的鳥類常在低海拔山區活動,牠們敏捷的行動和靈活的捕食技巧使牠們成為蛙類的潛在威脅。
黑冠麻鷺(Gorsachius melanolophus)是另一種在台灣常見的夜行性鷺科鳥類。這種鳥類喜歡在森林邊緣或溪流附近活動,以各種水生動物為食,包括蛙類。黑冠麻鷺的夜行性使牠們能夠捕食那些在夜間活動的蛙類,這些蛙類可能在白天較少受到其他鳥類的威脅。
魚鷹(Pandion haliaetus)雖然主要以魚類為食,但在某些情況下也會捕食大型的蛙類。這種猛禽在台灣主要是冬候鳥,牠們強大的飛行能力和銳利的爪子使牠們能夠從水面上抓取獵物。雖然捕食蛙類不是魚鷹的主要行為,但這種情況的發生反映了生態系統中食物網的複雜性。
尖尾鴨(Anas acuta)是一種在台灣常見的候鳥,雖然主要以植物性食物為主,但也會捕食小型動物,包括蝌蚪和小型蛙類。這種行為特別在繁殖季節明顯,當牠們需要更多蛋白質來支持繁殖活動時。
大卷尾(Dicrurus macrocercus)雖然主要以昆蟲為食,但也被觀察到會捕食小型蛙類。這種行為通常發生在昆蟲數量減少的季節,反映了鳥類在食物資源有限時的適應能力。
這些鳥類對蛙類的捕食行為不僅影響蛙類的種群動態,還在更大的生態系統中扮演著重要角色。例如,牠們可能通過捕食來控制某些優勢蛙類的數量,從而為其他蛙類創造生存空間,維持蛙類群落的多樣性。同時,這些鳥類也可能成為蛙類毒素或疾病的載體,在不同水體或棲息地之間傳播。
然而,隨著人類活動的增加,這些鳥類和牠們的蛙類獵物都面臨著棲息地喪失和環境污染等威脅。例如,濕地的減少和水質污染不僅直接影響蛙類的生存,也減少了這些捕食鳥類的覓食場所。因此,保護這些鳥類及其棲息地不僅對鳥類本身重要,也對維持整個生態系統的平衡至關重要。
了解台灣常見吃蛙類的鳥類不僅有助於我們更好地理解當地的生態系統動態,還為生態保護和管理提供了重要的科學依據。通過保護這些鳥類和牠們的棲息地,我們實際上是在保護一個更大、更複雜的生態網絡,其中包括了多樣的蛙類種群以及與之相關的其他生物。
9.2 蛙類生存競爭
蛙類生存競爭是生態學中一個複雜而引人入勝的主題,反映了自然界中物種為了有限資源而進行的持續鬥爭。在蛙類群落中,競爭主要發生在食物、棲息地、繁殖場所等資源的爭奪上,這種競爭不僅影響個體的生存和繁衍,還塑造了整個生態系統的結構和功能。
食物競爭是蛙類之間最常見的競爭形式之一。雖然不同種類的蛙類可能有不同的食性偏好,但在資源有限的情況下,它們常常會爭奪相同的食物來源。例如,在台灣的水稻田生態系統中,澤蛙(Fejervarya limnocharis)和黑眶蟾蜍(Duttaphrynus melanostictus)可能會競爭相似的昆蟲獵物。這種競爭可能導致一些物種改變其覓食行為或時間,以減少直接競爭。
棲息地競爭是另一個重要的競爭形式。不同的蛙類可能偏好相似的微棲息地,如特定的水體類型或植被結構。在台灣的山區溪流中,盤古蟾蜍(Bufo bankorensis)和梭德氏赤蛙(Rana sauteri)可能會競爭相同的溪流邊緣棲息地。這種競爭可能導致某些物種被迫使用次優的棲息地,從而影響其生存和繁殖成功率。
繁殖場所的競爭在蛙類生活史中扮演著關鍵角色。許多蛙類需要特定的水體環境來產卵和孵化蝌蚪。在台灣的低海拔地區,如虎皮蛙(Hoplobatrachus rugulosus)和澤蛙可能會競爭相同的繁殖水體。這種競爭不僅涉及空間的爭奪,還可能包括聲學干擾,即雄性個體通過鳴叫來爭奪配偶和領地。
聲學干擾是一種特殊的競爭形式,在蛙類中尤為重要。不同種類的蛙類可能會通過調整其鳴叫的頻率、強度或時間來減少相互干擾。例如,在台灣的樹蛙科中,褐樹蛙(Buergeria robusta)和莫氏樹蛙(Kurixalus moltrechti)可能會通過錯開鳴叫時間來減少競爭。
種間競爭的結果可能導致生態位分化(niche differentiation)。長期的競爭壓力可能促使不同種類的蛙類發展出特化的形態、行為或生理特徵,以更好地利用特定的資源。例如,在台灣的森林生態系統中,不同種類的樹蛙可能會特化於不同高度的樹冠層,以減少直接競爭。
入侵種的引入可能加劇蛙類之間的競爭。在台灣,外來的福建大頭蛙(Kaloula pulchra)的引入可能對本地蛙類造成競爭壓力,尤其是在都市化地區的有限水體中。這種競爭可能導致本地種的數量減少或分布範圍縮小。
氣候變化也可能改變蛙類之間的競爭動態。隨著氣溫升高,某些適應較冷環境的蛙類可能被迫向更高海拔遷移,這可能導致它們與原本生活在高海拔的物種產生新的競爭關係。例如,台灣的山椒魚(Hynobius formosanus)可能面臨來自低海拔遷移上來的蛙類的競爭壓力。
人類活動對蛙類競爭的影響不容忽視。棲息地破壞和片段化可能加劇競爭,因為可用的資源變得更加有限。例如,都市化過程中,水泥化的溝渠可能成為多種蛙類唯一可用的繁殖場所,導致競爭加劇。
蛙類之間的競爭並非總是負面的。適度的競爭可能促進物種的共存和多樣性。例如,通過資源分割(resource partitioning),不同種類的蛙類可以在同一生態系統中共存,每種蛙類專門利用特定的資源或微棲息地。
在研究蛙類競爭時,科學家們發現了一些有趣的現象。例如,某些蛙類表現出對競爭者的認知能力。研究表明,一些蛙類能夠根據競爭對手的存在來調整自己的行為,如改變鳴叫模式或避開特定的微棲息地。
競爭也可能隨著生活史階段的不同而變化。蝌蚪階段的競爭可能與成體階段大不相同。例如,在台灣的水域生態系統中,不同種類的蝌蚪可能競爭食物和空間,而成體則可能更多地競爭陸地棲息地和繁殖場所。
了解蛙類的競爭動態對於保護生物學和生態系統管理具有重要意義。通過了解這些複雜的相互作用,我們可以更好地預測環境變化對蛙類群落的影響,並制定更有效的保護策略。例如,在設計保護區時,考慮不同蛙類的競爭需求可以幫助創造更穩定和多樣化的生態系統。
此外,蛙類競爭的研究還為我們理解更廣泛的生態學原理提供了寶貴的見解。通過研究蛙類如何在競爭中共存,我們可以更好地理解生物多樣性的維持機制和群落結構的形成過程。這些知識不僅對於蛙類保護至關重要,還可以應用於其他生態系統的管理和恢復。
9.3 蛙類互利關係
蛙類互利關係是生態學中一個令人著迷的領域,揭示了這些兩棲動物如何與其他生物形成互惠共生的關係。這種關係不僅涉及蛙類與其他物種之間的直接互動,還包括了更廣泛的生態系統功能。蛙類的互利關係展現了自然界中複雜而精妙的平衡,也為我們理解生態系統的運作提供了重要視角。
一個典型的蛙類互利關係例子是某些樹蛙與植物之間的共生。在熱帶雨林中,一些樹蛙物種利用特定植物的葉子作為產卵和育幼的場所。這些葉子通常會形成一個天然的水槽,為蝌蚪提供安全的生長環境。作為回報,蛙類的排泄物和死亡的蝌蚪為植物提供了豐富的養分。這種關係在台灣的山區森林中也可以觀察到,例如莫氏樹蛙(Kurixalus moltrechti)經常在某些大型葉片植物上產卵。
另一個有趣的互利關係發生在某些蛙類和真菌之間。一些研究發現,某些真菌物種能夠生長在蛙類的皮膚上,這些真菌能夠產生抗菌物質,幫助蛙類抵抗皮膚病原體,特別是對抗致命的壺菌病(chytridiomycosis)。反過來,蛙類的皮膚為這些有益真菌提供了理想的生長環境。這種關係對於蛙類的健康和生存至關重要,尤其在面對全球兩棲類危機時。
蛙類與某些昆蟲之間也存在互利關係。例如,一些蛙類物種會選擇在蟻窩或白蟻窩附近生活。蛙類可以捕食接近窩穴的小型昆蟲,同時蟻類或白蟻的存在也為蛙類提供了某種程度的保護,因為許多潛在捕食者會避開這些具有攻擊性的昆蟲群落。這種關係在台灣的低海拔地區也有觀察到,如麗琉椿象(Cantao ocellatus)與某些地棲蛙類的關係。
一些蛙類與微生物之間也形成了互利關係。蛙類皮膚上的細菌群落不僅幫助維持皮膚健康,還可能在防禦病原體方面發揮作用。這些微生物可能產生抗菌物質,幫助宿主抵抗感染。同時,蛙類的皮膚分泌物為這些有益細菌提供了理想的生存環境。這種關係在面對全球兩棲類疾病威脅時顯得尤為重要。
在某些生態系統中,蛙類與魚類之間也可能形成互利關係。例如,在一些淡水環境中,某些蛙類的蝌蚪可能會清理魚類身上的寄生蟲或死皮。這種行為不僅為蝌蚪提供了食物來源,還幫助魚類保持健康。雖然這種關係在台灣尚未有明確記錄,但在其他地區的研究可以為我們理解類似的生態互動提供參考。
蛙類與某些鳥類之間的關係也可能存在互利性質。雖然許多鳥類捕食蛙類,但在某些情況下,蛙類的存在可能吸引昆蟲,間接為某些鳥類提供食物來源。同時,鳥類的活動可能驚擾昆蟲,使其更容易被蛙類捕獲。這種複雜的互動在台灣的濕地生態系統中可能存在,但需要更多研究來確認。
在農業生態系統中,蛙類與人類之間也形成了一種特殊的互利關係。蛙類通過捕食農業害蟲來幫助農民控制害蟲數量,而農民則通過維護適合蛙類生存的環境(如保留水田、減少農藥使用)來支持蛙類種群。在台灣的有機水稻田中,這種關係尤為明顯,澤蛙(Fejervarya limnocharis)等物種在害蟲控制中發揮了重要作用。
某些蛙類與水生植物之間也存在互利關係。蛙類可能幫助植物傳播花粉或種子,特別是那些在水邊活動的物種。同時,這些植物為蛙類提供了隱蔽和產卵的場所。在台灣的淡水濕地中,如台北赤蛙(Rana taipehensis)可能與某些水生植物之間存在這種關係。
蛙類的互利關係還體現在它們與整個生態系統之間的相互作用上。通過其捕食行為和作為其他動物的食物來源,蛙類在維持生態平衡中扮演著關鍵角色。它們參與能量流動和物質循環,對維持生態系統的健康和穩定性至關重要。
研究蛙類的互利關係不僅有助於我們更好地理解生態系統的複雜性,還為保護生物學提供了重要的洞見。通過了解這些關係,我們可以制定更全面的保護策略,不僅關注蛙類本身,還要考慮到與之相關的其他生物。
此外,蛙類互利關係的研究還可能為生物技術和醫學領域提供靈感。例如,從蛙類皮膚微生物群落中分離出的抗菌物質可能成為新型抗生素的來源。同樣,蛙類與真菌的共生關係也可能為人類對抗真菌感染提供新的思路。
在面對全球環境變化的背景下,理解蛙類的互利關係變得尤為重要。氣候變化、棲息地破壞和污染等因素可能擾亂這些精妙的生態關係。通過深入研究這些互利關係,我們可以更好地預測環境變化對生態系統的影響,並制定更有效的保護措施。
總的來說,蛙類互利關係的研究不僅豐富了我們對自然界複雜性的理解,還為生態保護、農業實踐和生物技術的發展提供了重要的理論基礎和實際應用前景。這個領域的持續探索無疑將為我們揭示更多自然界的奧秘,並為人類社會帶來潛在的益處。
9.4 蛙類作為生物指標
蛙類作為生物指標在生態學和環境科學中扮演著極其重要的角色。這些兩棲動物對環境變化高度敏感,使它們成為評估生態系統健康狀況的理想指標生物。蛙類的生理特性、生活史特徵以及其在生態系統中的獨特位置,使它們能夠反映出環境質量的細微變化,從而為科學家和環境管理者提供寶貴的信息。
蛙類作為生物指標的一個主要原因是它們具有滲透性皮膚。這種特殊的皮膚結構使蛙類極易吸收水中和空氣中的污染物。例如,在台灣的水稻田生態系統中,澤蛙(Fejervarya limnocharis)常被用來監測農藥污染。研究人員通過分析澤蛙體內的農藥殘留量,可以評估農業活動對環境的影響程度。這種方法不僅能夠檢測出水體中的污染物,還能反映出這些污染物在食物鏈中的生物累積效應。
蛙類的生活史特徵也使其成為優秀的生物指標。大多數蛙類在幼體階段生活在水中,成體則在陸地上活動。這種跨界的生活方式使蛙類能夠同時反映水陸兩種環境的狀況。在台灣的山區溪流中,盤古蟾蜍(Bufo bankorensis)的數量變化可以反映出溪流水質和周邊陸地環境的變化。如果盤古蟾蜍的種群數量出現異常波動,可能預示著溪流生態系統正面臨威脅。
蛙類對環境pH值的變化特別敏感,這使它們成為酸雨和水體酸化的重要指標。在台灣北部的山區,如觀霧地區,研究人員通過監測當地蛙類種群的變化來評估酸雨對森林生態系統的影響。如果某些酸敏感的蛙類物種,如莫氏樹蛙(Kurixalus moltrechti),出現數量減少或分布範圍縮小的情況,可能意味著該地區正遭受酸雨的侵襲。
蛙類的鳴叫行為也是一個重要的指標。不同物種的蛙類有特定的鳴叫時間和頻率,這些聲音信號可以反映出環境條件的變化。例如,在台灣的低海拔地區,褐樹蛙(Buergeria robusta)的鳴叫模式變化可能暗示著氣候異常或棲息地干擾。通過長期監測蛙類的鳴叫活動,科學家們可以追蹤環境變化的趨勢。
蛙類的畸形率也是一個重要的生物指標。環境中的某些污染物,如重金屬或內分泌干擾物,可能導致蛙類出現畸形。在台灣的工業區周邊水域,研究人員通過觀察當地蛙類的畸形率來評估工業污染的程度。高畸形率可能意味著環境中存在嚴重的化學污染。
蛙類對氣候變化的反應也使其成為全球變暖的重要指標。在台灣,隨著氣溫升高,一些原本生活在低海拔地區的蛙類物種開始向高海拔遷移。例如,台北赤蛙(Rana taipehensis)的分布範圍變化可能反映出局部氣候的變化趨勢。通過長期監測蛙類的分布變化,科學家們可以更好地理解並預測氣候變化對生態系統的影響。
蛙類作為生物指標還體現在它們對棲息地破壞的敏感性上。由於許多蛙類物種對特定的微棲息地有嚴格的要求,棲息地的輕微變化都可能導致蛙類種群的顯著波動。在台灣的都市化地區,諸如台北樹蛙(Rhacophorus taipeianus)等對環境要求較高的物種的消失可能預示著棲息地質量的下降。
蛙類的遺傳多樣性也是一個重要的指標。環境壓力可能導致蛙類種群的遺傳多樣性下降,這反過來又會降低種群應對環境變化的能力。在台灣的一些孤立的山區種群中,如觀霧山椒魚(Hynobius fuca)的遺傳多樣性變化可以反映出棲息地破碎化和氣候變化的影響。
蛙類作為食物網中的關鍵成員,其種群動態可以反映整個生態系統的健康狀況。例如,在台灣的水田生態系統中,蛙類的數量變化可能影響依賴它們為食的鳥類和蛇類的種群,進而影響整個食物網的結構。因此,通過監測蛙類種群,我們可以洞察更廣泛的生態變化。
然而,使用蛙類作為生物指標也面臨一些挑戰。首先,蛙類種群的自然波動可能掩蓋環境變化造成的影響,這需要長期和系統的監測來區分。其次,不同物種的蛙類對環境因素的敏感度不同,選擇合適的指標物種至關重要。最後,蛙類的生物指標作用可能受到多種因素的共同影響,這要求研究者謹慎解釋監測結果。
儘管如此,蛙類作為生物指標的價值仍然是不可替代的。它們提供了一種相對簡單、成本效益高的方法來評估環境質量和生態系統健康。通過持續監測蛙類種群的變化,我們可以及早發現潛在的環境問題,並採取相應的保護措施。
在台灣,蛙類作為生物指標的應用已經在多個領域展開。從濕地生態系統的評估到城市化影響的研究,從農業污染的監測到氣候變化的追蹤,蛙類都發揮了重要的指示作用。這些研究不僅豐富了我們對本地生態系統的理解,還為制定有效的環境政策和保護策略提供了科學依據。
9.5 蛙類在食物網中的地位
蛙類在生態系統的食物網中扮演著關鍵的角色,它們既是捕食者,也是被捕食的對象,因此在能量傳遞和生態平衡中發揮著重要作用。作為捕食者,蛙類主要以昆蟲和其他小型無脊椎動物為食,包括蚊子、蒼蠅、蛾子、甲蟲等。某些大型蛙類甚至能捕食小型哺乳動物、鳥類和其他兩棲類。這種捕食行為不僅幫助控制昆蟲種群數量,還維持了生態系統的平衡。
在水生環境中,蝌蚪階段的蛙類主要以藻類和有機碎屑為食,在水域生態系統中扮演著初級消費者的角色。隨著蝌蚪變態為成蛙,其食性也隨之改變,成為陸地和水生環境中的次級消費者。這種生活史階段的轉變使得蛙類能夠在不同的生態位中發揮作用,連接水陸生態系統。
作為被捕食者,蛙類是許多動物的重要食物來源。水生階段的蝌蚪常被魚類、水生昆蟲和某些鳥類捕食。成蛙則面臨來自多種捕食者的威脅,如蛇類、鳥類(如鷺鷥和鷹)、哺乳動物(如浣熊和水獺)以及大型魚類。在某些地區,蛙類甚至成為人類的食物來源。這種被捕食的角色使得蛙類成為將初級生產者的能量傳遞到更高營養級別的重要媒介。
蛙類的存在對於維持濕地和水域生態系統的健康至關重要。它們通過控制昆蟲種群數量來幫助維持生態平衡,同時又為其他動物提供食物來源。例如,在某些濕地生態系統中,蛙類可以控制蚊子的數量,從而間接影響疾病的傳播。另一方面,蛙類數量的變化可能會影響依賴它們為食的捕食者種群,如某些鳥類和爬行動物。
蛙類在食物網中的位置也反映了生態系統的健康狀況。由於蛙類對環境變化特別敏感,它們常被視為生態系統健康的指標物種。水質污染、棲息地破壞或氣候變化等因素都可能通過影響蛙類種群而對整個食物網造成連鎖反應。例如,農藥的使用可能直接毒害蛙類,或通過減少其食物來源間接影響蛙類種群,進而影響依賴蛙類為食的捕食者。
在某些生態系統中,蛙類還可能影響營養循環。它們的排泄物可以為水生植物和微生物提供養分,促進初級生產。當蛙類在水陸之間遷移時,它們還可能將養分從水生環境帶到陸地環境,或反之,從而在不同生態系統之間形成養分流動。
蛙類在食物網中的作用也因地區和物種而異。例如,在熱帶雨林中,某些樹蛙可能主要以樹冠層的昆蟲為食,而在溫帶地區的池塘中,水蛙可能更多地捕食水生昆蟲和小魚。這種多樣性使得蛙類能夠適應不同的生態位,在各種生態系統中發揮作用。
此外,蛙類的季節性行為也影響其在食物網中的角色。在繁殖季節,大量蛙類聚集在水體周圍,可能會吸引更多捕食者,形成短暫但重要的食物資源集中地。相反,在非繁殖季節,蛙類可能分散到更廣闊的區域,其在食物網中的影響也隨之分散。
值得注意的是,全球範圍內的兩棲類衰退現象對食物網結構產生了深遠影響。蛙類種群的減少不僅直接影響依賴它們為食的動物,還可能導致其捕食對象(如某些昆蟲)的種群激增,進而影響植被和其他生態系統組成部分。這種連鎖反應凸顯了蛙類在維持生態平衡中的重要性,也強調了保護蛙類及其棲息地的必要性。
10. 蛙類種群生態學
蛙類種群生態學是研究蛙類種群動態、結構和相互作用的學科。這一領域關注蛙類種群如何隨時間和空間變化,以及影響這些變化的因素。蛙類種群的大小和分布受到多種因素的影響,包括出生率、死亡率、遷入和遷出。
蛙類種群結構通常呈現年齡和大小的分層,這反映了不同年齡階段的個體在種群中的比例。例如,在某些蛙類種群中,可能會觀察到大量的幼蛙和較少的成年個體,這種結構可能與高死亡率或特定的生活史策略有關。
蛙類的遷徙行為對種群動態有重要影響。許多蛙類物種每年都會在繁殖地和非繁殖地之間進行季節性遷徙。這種行為不僅影響局部種群的大小,還促進了基因流動,有助於維持種群的遺傳多樣性。
種群間的相互作用,特別是形成大的族群系統(metapopulation),是蛙類種群生態學的另一個重要方面。在這種系統中,分散的局部種群通過個體的遷移和基因交流而相互聯繫。這種結構增加了整體種群的穩定性和適應能力。
監測蛙類種群是理解其生態學和制定保護策略的關鍵。常用的監測方法包括標記-重捕法、鳴叫調查和目視計數等。這些方法有助於評估種群大小、密度和長期趨勢,為保護決策提供重要依據。
蛙類種群生態學的研究對於保護工作至關重要。通過了解影響蛙類種群的因素,科學家們可以預測潛在的威脅並制定有效的保護策略,確保這些重要的兩棲動物在生態系統中繼續發揮其關鍵作用。
10.1 蛙類種群結構
蛙類種群結構是蛙類種群生態學中的核心概念,它描述了一個種群內部的組成和特徵。這個結構包含了多個重要的維度,如年齡分布、性別比例、體型大小分布以及遺傳多樣性等。這些因素共同塑造了蛙類種群的整體特徵,並深刻影響著種群的動態和演化潛力。
年齡結構是理解蛙類種群的關鍵要素。一個典型的蛙類種群通常包括不同年齡階段的個體,從剛變態的幼蛙到成年繁殖個體。這種年齡分層反映了種群的繁殖成功率和生存狀況。例如,在一個健康的種群中,我們可能會觀察到大量的幼蛙和亞成體,以及相對較少但穩定的成年個體。這種結構表明種群有良好的補充和生存率。相反,如果一個種群缺乏年輕個體,可能意味著繁殖失敗或幼體存活率低,這可能預示著種群面臨衰退的風險。
性別比例是另一個重要的種群結構指標。在理想情況下,大多數蛙類種群的性別比例應接近1:1。然而,環境因素、選擇性捕食或人為干擾可能導致性別比例失衡。例如,某些環境污染物可能影響蝌蚪的性別分化,導致雄性或雌性個體比例異常增高。性別比例的失衡可能嚴重影響種群的繁殖潛力和長期生存能力。
體型大小分布是蛙類種群結構的另一個重要方面。在同一種群中,個體大小可能因年齡、性別和環境條件而異。通常,雌性蛙類比雄性個體更大,這與它們需要產生大量卵子有關。體型大小分布可以反映種群的營養狀況、生長率和競爭壓力。例如,如果一個種群中缺乏大型個體,可能表明環境資源匱乏或面臨高度捕食壓力。
遺傳多樣性是種群結構的一個不可忽視的維度,儘管它不像年齡或大小那樣直觀可見。高度的遺傳多樣性通常意味著種群具有更強的適應能力和更好的長期生存前景。相反,遺傳多樣性低的種群可能更容易受到環境變化和疾病的影響。例如,在面臨新的病原體時,遺傳多樣性高的種群可能有更多的個體能夠產生有效的免疫反應。
蛙類種群的空間分布也是種群結構的重要組成部分。許多蛙類物種表現出聚集分布的特徵,特別是在繁殖季節。這種分布模式可能導致局部種群密度高度變化,進而影響個體間的競爭和繁殖機會。例如,在一個池塘生態系統中,蛙類可能集中在特定的微棲息地,如水生植物豐富的區域,而在其他區域則較為稀疏。
季節性變化是影響蛙類種群結構的另一個重要因素。許多蛙類物種在非繁殖季節可能分散在較大的區域內,而在繁殖季節則聚集在特定的水體周圍。這種季節性聚集不僅改變了局部種群的密度和結構,還可能影響捕食壓力和資源競爭的強度。
種群結構還受到密度依賴效應的影響。當種群密度增加時,個體間的競爭加劇,可能導致生長率下降、死亡率上升或繁殖成功率降低。這種密度依賴效應可能會改變種群的年齡結構和體型分布。例如,在高密度條件下,幼體的生存率可能下降,導致種群年齡結構向成年個體傾斜。
外來種的入侵也可能顯著改變本地蛙類的種群結構。入侵種可能通過競爭、捕食或引入新疾病來影響本地種群。例如,入侵的牛蛙(Rana catesbeiana)不僅直接捕食小型本地蛙類,還可能佔據它們的棲息地,從而改變本地種群的年齡結構和空間分布。
人類活動對蛙類種群結構的影響不容忽視。棲息地破壞、污染和氣候變化等因素可能導致種群結構的顯著變化。例如,農藥的使用可能對幼體和亞成體造成更大的影響,從而改變種群的年齡結構。城市化過程中的棲息地碎片化可能導致小而孤立的亞種群,增加近親繁殖的風險,降低遺傳多樣性。
了解蛙類種群結構對於制定有效的保護策略至關重要。通過監測種群結構的變化,研究人員可以及早發現潛在的問題並採取相應的保護措施。例如,如果發現某個種群缺乏年輕個體,可能需要重點保護繁殖棲息地或改善幼體的生存環境。同樣,如果發現種群的遺傳多樣性降低,可能需要考慮引入其他種群的個體以增加基因流動。
在研究蛙類種群結構時,科學家們運用多種方法和技術。標記-重捕法是一種常用的技術,可以提供關於種群大小、生存率和個體生長的信息。聲學監測技術則可以用來估計繁殖季節的雄性數量和分布。最近,分子生物学技術的應用,如DNA分析,為研究種群的遺傳結構和基因流動提供了新的工具。
綜上所述,蛙類種群結構是一個多維度、動態變化的概念,它反映了種群的當前狀態和未來潛力。通過深入研究和長期監測蛙類種群結構,我們不僅可以更好地理解這些生物的生態學特徵,還可以為其保護提供科學依據,確保蛙類在生態系統中持續發揮其重要作用。
10.2 蛙類擴散與遷徙
蛙類的擴散與遷徙是其生活史中的重要行為,對種群動態、基因流動和生態系統功能有著深遠的影響。這些行為模式不僅反映了蛙類對環境變化的適應,也展示了它們在生態系統中的重要角色。
擴散通常指個體從出生地或繁殖地向外移動的過程。對蛙類而言,擴散行為主要發生在變態後的幼蛙階段。這個時期的蛙類離開水域,開始向周圍陸地環境擴散。擴散行為有助於減少同一地點的種內競爭,同時也促進了種群的基因交流。例如,在某些樹蛙物種中,剛變態的幼蛙可能會在幾天內迅速擴散到周圍數百米甚至更遠的區域。這種擴散模式不僅降低了局部種群的密度依賴效應,也有助於物種的地理分布擴張。
遷徙則通常指蛙類在不同季節或生活階段之間的大規模移動。最典型的遷徙行為是成年蛙類在繁殖季節向水體遷移。許多蛙類物種在非繁殖季節生活在陸地環境中,但在繁殖季節會集體遷徙到特定的水體進行繁殖。這種遷徙行為可能涉及相當長的距離,有些物種甚至能夠跨越數公里的陸地到達繁殖地。例如,北美的木蛙(Rana sylvatica)每年春季都會從越冬地遷徙到繁殖池塘,這個過程中它們可能需要穿越森林、草地甚至是人類居住區。
蛙類的擴散和遷徙行為受多種因素影響。環境因素如溫度、濕度和降雨量往往是觸發這些行為的主要原因。例如,許多蛙類物種會在雨季開始時開始遷徙,因為這時水體充足,適合繁殖。此外,光週期的變化也可能作為遷徙的信號。內部因素如激素水平的變化也在調節這些行為中發揮重要作用。
地形特徵對蛙類的擴散和遷徙路線有顯著影響。蛙類通常會選擇濕度較高的路徑進行移動,如溪流沿岸或低窪地區。地形障礙如山脈或大型水體可能阻礙某些物種的擴散,而對其他物種則可能提供擴散通道。例如,河谷系統可能成為某些蛙類物種沿著其擴散的主要路徑。
人類活動對蛙類的擴散和遷徙造成了顯著影響。棲息地破碎化是一個主要問題,道路、城市發展和農業活動可能切斷蛙類的遷徙路線或阻礙其擴散。例如,在許多地區,每年都有大量蛙類在遷徙過程中被汽車碾壓。為了減少這種影響,一些地方建立了蛙類通道或在繁殖季節臨時關閉某些道路。
氣候變化也正在改變蛙類的擴散和遷徙模式。隨著氣溫升高,一些蛙類物種的分布範圍正在向高緯度或高海拔地區擴展。這種範圍的變化可能導致新的種間競爭或捕食關係,進而影響整個生態系統的平衡。
蛙類的擴散和遷徙行為對維持種群的遺傳多樣性至關重要。通過這些行為,不同亞種群之間能夠進行基因交流,減少近親繁殖的風險,增加種群的適應能力。然而,如果這些行為受到阻礙,可能導致種群的遺傳隔離和局部滅絕。
研究人員使用多種方法來研究蛙類的擴散和遷徙行為。傳統的標記-重捕法可以提供關於個體移動距離和方向的信息。近年來,無線電遙測技術的應用使得研究人員能夠更精確地追蹤個體的移動路徑。此外,基因標記技術也被廣泛應用於研究種群間的基因流動和擴散模式。
蛙類的擴散和遷徙行為在生態系統中扮演著重要角色。通過這些行為,蛙類能夠在不同棲息地之間轉移能量和物質,連接陸地和水域生態系統。例如,當成年蛙類從陸地遷徙到水體進行繁殖時,它們將陸地生態系統中獲得的能量帶入水域生態系統,這種能量轉移對維持水域生態系統的平衡具有重要意義。
某些蛙類物種展現出獨特的擴散和遷徙策略。例如,一些樹蛙物種能夠利用樹冠層進行長距離擴散,這種策略使它們能夠跨越地面上的障礙物。另一方面,一些生活在乾旱環境中的蛙類物種則採取保守的策略,它們可能一生都停留在同一個水坑附近,只在雨季時進行短距離的移動。
了解蛙類的擴散和遷徙行為對於制定有效的保護策略至關重要。通過識別關鍵的遷徙路線和擴散廊道,保護工作者可以優先保護這些區域,確保蛙類種群的連通性。例如,在道路建設過程中,可以設計並建造專門的蛙類通道,以減少道路對蛙類遷徙的阻礙。
蛙類的擴散和遷徙行為還為我們理解生態系統的動態提供了重要窗口。通過研究這些行為,科學家們可以更好地理解氣候變化、棲息地破壞和其他環境變化對生物多樣性的影響。這些研究不僅有助於保護蛙類,還為整個生態系統的管理和保護提供了重要依據。
綜上所述,蛙類的擴散與遷徙是一個複雜而重要的生態過程,它涉及多個尺度的空間和時間動態,反映了蛙類對環境的適應以及在生態系統中的重要作用。深入理解這些行為不僅能增進我們對蛙類生態學的認知,還能為生物多樣性保護和生態系統管理提供重要的科學基礎。
10.3 蛙類種群間動態
蛙類種群間動態,也稱為族群系統動態,是研究由多個局部種群組成的大尺度種群結構及其變化的重要領域。這一概念對於理解蛙類在景觀尺度上的分布、持續性和遺傳多樣性維持至關重要。族群系統模型描述了一系列分散但相互連接的局部種群,這些種群通過個體的遷移和基因流動而相互影響。
在蛙類中,族群系統結構普遍存在,特別是在棲息地呈現斑塊狀分布的情況下。例如,在一個由多個池塘組成的濕地系統中,每個池塘可能支持一個局部種群,這些種群通過蛙類的擴散和遷徙行為而相互連接,形成一個大的族群系統。這種結構使得即使某些局部種群因環境變化或隨機事件而滅絕,整個系統仍能通過其他種群的個體重新殖民而維持。
族群系統動態的核心是局部種群之間的滅絕和重新殖民過程。在蛙類中,這一過程尤為明顯。例如,某些臨時性水體中的蛙類種群可能在乾旱年份完全消失,但在雨量充沛的年份又被鄰近種群的個體重新殖民。這種動態平衡對於維持整個族群系統的長期穩定性至關重要。
影響蛙類族群系統動態的因素眾多,包括棲息地的質量和分布、棲息地之間的連通性、環境波動以及人類活動等。棲息地質量直接影響局部種群的生存和繁殖成功率。高質量的棲息地可能成為整個系統的「源」,持續向周圍低質量棲息地輸出個體。相反,低質量棲息地可能成為「匯」,需要不斷從其他種群獲得補充才能維持。
棲息地之間的連通性是維持族群系統功能的關鍵。對蛙類而言,連通性不僅指地理距離,還包括棲息地之間的景觀特徵。例如,一條溪流或濕地走廊可能大大增強相距較遠的兩個池塘之間的連通性,而一條繁忙的公路則可能成為阻隔近距離棲息地之間基因流動的屏障。
環境波動對蛙類族群系統動態有顯著影響。季節性變化、極端天氣事件如乾旱或洪水,都可能導致局部種群的波動甚至滅絕。然而,族群系統結構提供了一種緩衝機制,使得整個系統能夠在面對局部擾動時保持穩定。例如,在一個由多個池塘組成的系統中,即使某些池塘因乾旱而乾涸,其他仍有水的池塘可以作為種群的避難所,待條件改善後再重新擴散。
人類活動對蛙類族群系統動態的影響不容忽視。棲息地破壞和破碎化是最直接的威脅,它們不僅減少了可用的棲息地面積,還降低了棲息地之間的連通性。例如,城市化過程中的道路建設和土地開發可能將原本連續的濕地系統分割成孤立的小斑塊,大大降低了蛙類在斑塊間移動的能力,從而影響整個族群系統的動態。
氣候變化也正在改變蛙類族群系統的動態。氣溫升高和降雨模式改變可能導致某些棲息地變得不適宜,迫使蛙類向更適宜的區域遷移。這種大尺度的分布變化可能重塑整個族群系統的結構,創造新的連接或切斷現有的連接。
在研究蛙類族群系統動態時,科學家們運用了多種方法和技術。標記-重捕法可以提供關於個體在不同局部種群之間移動的直接證據。遺傳學方法,如微衛星DNA分析,則可以揭示長期的基因流動模式和種群間的遺傳關係。此外,地理信息系統(GIS)技術的應用使得研究人員能夠在景觀尺度上分析棲息地分布和連通性。
族群系統動態對蛙類的遺傳多樣性維持有重要影響。通過個體在不同局部種群之間的移動,基因得以在更大範圍內流動,減少了近親繁殖的風險,增加了種群的適應潛力。然而,如果局部種群之間的連通性受到破壞,可能導致遺傳多樣性的喪失和適應能力的下降。
了解蛙類族群系統動態對制定有效的保護策略至關重要。傳統的保護方法往往集中於保護單個高質量棲息地,而族群系統的概念強調了保護棲息地網絡的重要性。例如,在設計保護區時,不僅要考慮包含高質量棲息地,還要確保這些棲息地之間有足夠的連通性,以維持整個系統的功能。
族群系統動態還為理解蛙類對環境變化的響應提供了新的視角。在面對氣候變化等大尺度環境變化時,具有健康族群系統結構的物種可能表現出更強的恢復力。這是因為它們可以通過局部種群之間的相互補充來緩衝環境波動的影響。
某些蛙類物種展現出特別有趣的族群系統動態。例如,一些生活在臨時性水體中的蛙類,其族群系統可能呈現高度的時空動態,局部種群的存在與消失與降雨模式密切相關。另一方面,一些森林樹蛙的族群系統可能更為穩定,但對森林破碎化特別敏感。
族群系統動態的研究不僅有助於我們理解蛙類的生態學,還為更广泛的生態理論提供了重要見解。例如,它為理解物種如何在異質性景觀中持續存在、如何應對環境變化以及如何維持遺傳多樣性提供了框架。這些見解對於其他類群的保護和管理同樣具有重要意義。
10.4 影響蛙類種群大小的因素
影響蛙類種群大小的因素是一個複雜而多元的議題,涉及生物學、生態學和環境科學的多個方面。這些因素可以broad分為自然因素和人為因素,它們相互作用,共同塑造了蛙類種群的動態變化。
首先,棲息地的可用性和質量是影響蛙類種群大小的最基本的因素之一。蛙類需要適宜的水域環境進行繁殖和幼體發育,同時也需要陸地環境用於覓食和冬眠。水體的pH值、溶氧量、污染程度等都直接影響蛙類的生存和繁殖。例如,酸化的水體會影響蛙卵的發育,而富營養化則可能導致水體缺氧,危及蝌蚪的生存。陸地棲息地的植被覆蓋、濕度和食物可用性也同樣重要。棲息地的減少或退化會直接導致種群大小的下降。
氣候因素,特別是溫度和降水,對蛙類種群大小有顯著影響。蛙類作為變溫動物,其活動和代謝強烈依賴於環境溫度。過高或過低的溫度都可能導致死亡率上升。降水模式影響水體的持續性和水量,進而影響繁殖成功率。例如,乾旱可能導致臨時性水體提前乾涸,造成大量蝌蚪死亡。氣候變化正在改變許多地區的溫度和降水模式,這對蛙類種群構成了新的挑戰。
食物可用性是另一個關鍵因素。蛙類主要以昆蟲和其他小型無脊椎動物為食。食物資源的豐富程度直接影響蛙類的生存、生長和繁殖。環境中昆蟲數量的波動可能導致蛙類種群的相應波動。例如,某些農業實踐可能減少昆蟲岐異度和豐富度,進而影響依賴這些昆蟲的蛙類種群。
捕食和競爭也是影響蛙類種群大小的重要生物因素。蛙類在不同生活階段面臨不同的捕食壓力,從魚類和水生昆蟲對蝌蚪的捕食,到蛇類、鳥類和哺乳動物對成蛙的捕食。捕食壓力的變化可能導致種群大小的顯著波動。同時,種間和種內競爭也會影響資源的可用性和個體的生存率。例如,在資源有限的情況下,高密度可能導致生長速度減慢和死亡率增加。
疾病是影響蛙類種群的另一個重要因素。近年來,壺菌病(Chytridiomycosis)成為全球蛙類面臨的主要威脅之一。這種由真菌引起的疾病已導致多個物種的種群崩潰甚至滅絕。其他病原體如病毒和寄生蟲也可能對種群造成重大影響。疾病的傳播和影響程度往往與環境條件和種群健康狀況密切相關。
人類活動對蛙類種群大小的影響尤為顯著。棲息地破壞和破碎化是最直接的威脅。城市化、農業擴張和森林砍伐不僅減少了可用棲息地的面積,還破壞了棲息地之間的連通性,影響種群之間的基因流動。水體污染,特別是農藥和化肥的使用,可能直接毒害蛙類或通過影響其食物鏈間接影響種群。例如,某些內分泌干擾物可能影響蛙類的性別分化和繁殖能力。
外來物種的入侵也是一個重要因素。入侵物種可能通過直接捕食、競爭資源或引入新的病原體來影響本地蛙類種群。例如,入侵的牛蛙不僅捕食小型本地蛙類,還可能傳播壺菌病。
氣候變化正在成為影響蛙類種群的一個越來越重要的因素。全球變暖不僅直接影響蛙類的生理過程,還可能改變其棲息地的適宜性。例如,某些高山物種可能因溫度上升而失去適宜棲息地。氣候變化還可能改變降水模式,影響繁殖水體的可用性和穩定性。
遺傳因素在決定蛙類種群大小和長期生存能力方面也扮演重要角色。小而孤立的種群可能面臨遺傳多樣性喪失和近親繁殖的風險,這可能降低其適應環境變化的能力。遺傳多樣性的維持對於種群應對疾病和環境變化至關重要。
人為干擾,如道路建設和交通,也可能對蛙類種群造成直接影響。道路不僅分割棲息地,還可能成為蛙類遷徙和擴散的障礙。道路死亡是許多蛙類種群面臨的重要威脅,特別是在繁殖季節的遷徙期間。
管理措施和保護政策的實施或缺失也會影響蛙類種群大小。有效的保護措施,如建立保護區、恢復退化棲息地、控制污染等,可以幫助維持或增加蛙類種群。相反,缺乏適當的管理可能導致種群持續下降。
理解這些影響蛙類種群大小的複雜因素對於制定有效的保護策略至關重要。它要求我們採取綜合的(comprehensive和integrativ)e的方法,同時考慮生態、環境和人類因素。只有充分了解這些因素及其相互作用,我們才能更好地預測和管理蛙類種群的變化,確保這些重要的兩棲動物在生態系統中持續發揮其關鍵作用。
10.5 蛙類種群監測
蛙類種群監測是生態學和保育生物學中的一個重要領域,它為理解蛙類種群動態、評估保育措施效果和制定管理策略提供了關鍵信息。這項工作涉及多種方法和技術,旨在準確估計種群大小、追蹤長期趨勢,並及時發現潛在的威脅。
聲學調查是蛙類種群監測中最常用的方法之一。由於大多數蛙類雄性在繁殖季節會發出特徵性的叫聲,研究人員可以通過記錄和分析這些叫聲來估計種群大小和分布。這種方法特別適用於夜間活動的物種。研究人員可以在固定的採樣點進行聽覺調查,記錄聽到的不同物種及其相對豐度。近年來,自動錄音設備的使用大大提高了這種方法的效率。這些設備可以長期放置在野外,連續記錄環境聲音,之後通過聲音識別軟件分析蛙類的叫聲。這不僅提高了數據收集的時間分辨率,還減少了人為偏差。
視覺調查是另一種常用的監測方法。這包括在特定區域內進行系統的搜索,記錄觀察到的個體數量和物種。這種方法適用於日間活動的物種或者在繁殖水體周圍進行調查。研究人員通常會沿著預設的穿越線或在固定的樣點進行調查,以確保採樣的一致性和可比性。視覺調查可以提供關於種群結構的重要信息,如成年個體和幼體的比例。
捕捉-標記-重捕法(Capture-Mark-Recapture,CMR)是一種更為精確但也更耗時的監測方法。這種方法涉及捕捉個體、進行標記(如剪趾法或植入微型晶片),然後釋放。在後續的採樣中,研究人員記錄重新捕獲的已標記個體和新捕獲的未標記個體的數量。通過統計模型,可以估算出總的種群大小。CMR方法不僅可以提供準確的種群估計,還可以提供關於個體生存率、生長率和移動模式的寶貴信息。
環境DNA (eDNA) 分析是近年來發展起來的一種創新監測技術。這種方法通過採集水樣或土壤樣本,分析其中存在的目標物種DNA來確定物種的存在。對於蛙類而言,這種方法特別有用,因為它們在水中繁殖和發育,會留下大量的DNA痕跡。eDNA技術的優勢在於它可以檢測到難以直接觀察的稀有或隱蔽物種,並且對環境干擾較小。
衛星遙感和地理信息系統(GIS)技術在大尺度蛙類種群監測中發揮著越來越重要的作用。這些技術可以幫助研究人員識別和監測適宜的蛙類棲息地,評估棲息地連通性,並預測氣候變化對蛙類分布的潛在影響。例如,研究人員可以使用衛星圖像跟蹤濕地面積的變化,或者利用地形數據模擬潛在的蛙類遷徙路線。
公民科學在蛙類種群監測中也扮演著重要角色。通過培訓志願者參與監測工作,研究人員可以在更大的空間和時間尺度上收集數據。許多國家和地區都建立了蛙類監測網絡,鼓勵公眾報告他們的蛙類觀察記錄。這些數據雖然可能存在一定的偏差,但在長期趨勢分析和罕見物種檢測方面非常有價值。
蛙類種群監測面臨著一些特殊挑戰。首先,許多蛙類物種的活動具有很強的季節性和天氣依賴性,這要求監測工作必須在適當的時間和條件下進行。其次,不同的監測方法可能對不同的物種或生活階段有不同的效果。例如,聲學調查可能低估雌性個體的數量,而視覺調查可能難以檢測到隱蔽性強的物種。因此,綜合使用多種監測方法通常是必要的。
長期監測對於理解蛙類種群動態至關重要。許多蛙類種群表現出強烈的年際波動,單次或短期的調查可能無法反映真實的種群趨勢。建立標準化的長期監測項目可以幫助研究人員區分自然波動和真正的種群下降趨勢。例如,北美兩棲類監測項目(North American Amphibian Monitoring Program)已經持續多年,為理解該地區蛙類種群變化提供了寶貴的數據。
新興的技術正在改變蛙類種群監測的方式。例如,環境基因組學技術可以通過分析水樣中的DNA不僅確定物種的存在,還能估計其相對豐度。人工智能和機器學習算法的應用正在提高聲音識別和圖像分析的效率和準確性。無人機技術則為難以到達的棲息地的監測提供了新的可能性。
蛙類種群監測的數據分析和解釋也是一個複雜的過程。研究人員需要考慮採樣誤差、檢測概率的變化以及環境因素的影響。高級統計模型,如占據模型(occupancy models)和N-mixture模型,正被廣泛用於處理這些複雜性,提供更準確的種群估計和趨勢分析。
監測數據的管理和共享也是蛙類種群監測中的重要環節。建立統一的數據庫,實現數據的標準化和開放共享,可以促進更廣泛的數據使用和跨區域的比較研究。例如,全球兩棲類評估(Global Amphibian Assessment)項目彙集了全球範圍內的兩棲類數據,為全球蛙類保育提供了重要的科學依據。
11. 蛙類棲息地與環境需求
蛙類的棲息地與環境需求是一個複雜而多樣的主題,反映了這些兩棲動物獨特的生活史特徵。蛙類需要同時適應水生和陸生環境,這使得它們對棲息地的要求尤為特殊。大多數蛙類物種需要清潔的水體進行繁殖和幼體發育,同時也需要適宜的陸地環境用於覓食和冬眠。
水體的特徵,如深度、水質、植被覆蓋等,直接影響蛙類的繁殖成功率。例如,某些物種偏好淺水區域產卵,而其他則需要較深的水體。水體的pH值、溶氧量和污染程度都是關鍵因素。陸地環境方面,蛙類需要適當的遮蔽物、濕度和食物來源。植被結構的複雜性對許多物種至關重要,提供了躲避捕食者和調節體溫的場所。
蛙類對環境溫度和濕度的要求也相當特殊,這些因素直接影響它們的生理功能和活動模式。某些物種能夠適應相對乾燥的環境,而others則需要持續的高濕度。氣候變化對蛙類棲息地的影響尤為顯著,改變了許多地區的溫度和降水模式,進而影響蛙類的分布和生存。
11.1 蛙類水生棲息地利用
蛙類對水生棲息地的利用是其生命週期中不可或缺的一部分,反映了這些兩棲動物獨特的生理需求和生態適應。水生環境對蛙類而言,不僅是繁殖和幼體發育的場所,也是許多成體賴以生存的重要棲息地。蛙類對水生棲息地的利用模式因物種、生活階段和季節而異,呈現出豐富多樣的生態學特徵。
對大多數蛙類來說,水體是繁殖活動的核心場所。在繁殖季節,成年蛙類會聚集在適宜的水體周圍。雄性通過鳴叫吸引雌性,而雌性則選擇合適的位置產卵。不同蛙類物種對產卵地點的選擇有著明顯的偏好。例如,有些物種喜歡在淺水區域產卵,這樣可以確保卵獲得充足的陽光和氧氣;而其他物種則可能選擇水深較深的區域,以減少被捕食的風險。水體的大小、深度、水質和植被覆蓋等因素都會影響蛙類的選擇。
蝌蚪階段是蛙類生命週期中最依賴水生環境的時期。不同物種的蝌蚪對水體環境有不同的適應和需求。有些蝌蚪主要生活在水體表層,以藻類和有機碎屑為食;而有些則更多地活動在水體底部。水體的特徵,如溫度、溶氧量、pH值和食物可用性,直接影響蝌蚪的生長和發育。例如,較高的水溫通常會加快蝌蚪的發育速度,但同時也可能增加死亡率。水體中的植被不僅為蝌蚪提供食物和遮蔽,還有助於維持水質。
成年蛙類對水生棲息地的利用模式更為複雜和多樣。有些物種,如水蛙屬(Rana)的許多成員,終生與水體密切相關,它們在水中覓食、躲避天敵,只在繁殖時才上岸。這些物種通常具有適應水生生活的特徵,如發達的蹼和流線型的身體。相比之下,有些蛙類物種雖然需要在水中繁殖,但成年後主要生活在陸地環境中,只在繁殖季節才返回水體。
水生植被在蛙類利用水生棲息地時扮演著關鍵角色。水生植物為蛙類提供了隱蔽場所,有助於躲避捕食者。同時,這些植物也是許多蛙類賴以生存的昆蟲和其他無脊椎動物的棲息地,為蛙類提供了豐富的食物來源。某些蛙類物種,如樹蛙科的一些成員,會利用水面上方的植被作為鳴叫和休息的場所。
水體的物理特性對蛙類的利用模式有重要影響。例如,水體的深度影響蛙類的活動範圍和捕食行為。淺水區域通常是許多物種的首選,因為這裡溫度較高,有利於體溫調節和新陳代謝。水流速度也是一個重要因素,一些適應流水環境的物種具有特殊的形態適應,如扁平的身體和吸盤狀的指端,使它們能夠在急流中生存。
季節性變化對蛙類利用水生棲息地的方式有顯著影響。在溫帶地區,許多蛙類物種會在冬季進入水底泥土中冬眠。這種行為要求水體不會完全結冰,並且底部有足夠的淤泥層。相反,在熱帶地區,一些蛙類物種可能在乾季時通過埋入濕潤的淤泥中來度過不利時期。
人類活動對蛙類水生棲息地的利用產生了深遠影響。水體污染、棲息地破壞和外來物種入侵是主要威脅。例如,農業逕流中的化肥和農藥可能導致水體富營養化和毒性物質積累,直接危害蛙類及其食物來源。水利工程,如水壩建設和河道整治,可能改變水文條件,破壞蛙類的繁殖場所。
氣候變化也正在改變蛙類對水生棲息地的利用模式。全球變暖導致的水溫升高和降水模式改變,可能影響水體的持續性和適宜性。例如,某些依賴臨時性水體繁殖的物種可能因為乾旱而失去繁殖場所。
為了應對這些挑戰,保護和恢復適宜的水生棲息地成為蛙類保育的關鍵。這包括維護自然的水文過程、控制污染、保護和恢復濱水植被等。一些地區通過建造人工濕地或改造現有水體,為蛙類創造適宜的棲息環境。這些努力不僅有助於保護蛙類,還能為整個水生生態系統帶來益處。
理解蛙類對水生棲息地的利用模式對於制定有效的保護策略至關重要。這要求研究人員全面考慮蛙類的生活史特徵、生態需求以及面臨的威脅。只有充分理解這些複雜的相互作用,我們才能確保蛙類及其賴以生存的水生生態系統的長期健康和穩定。
11.2 蛙類陸地棲息地利用
蛙類的陸地棲息地利用是其生態適應的重要組成部分,反映了這些兩棲動物在進化過程中對陸地環境的適應。儘管大多數蛙類物種仍然依賴水體進行繁殖,但它們在生命週期的大部分時間裡都在陸地環境中度過。蛙類對陸地棲息地的利用涉及多個方面,包括覓食、躲避天敵、調節體溫和季節性遷移等。
陸地環境為蛙類提供了豐富的食物來源。大多數成年蛙類是肉食性的,主要捕食各種昆蟲和其他小型無脊椎動物。不同蛙類物種根據自身的形態特徵和行為模式,在陸地環境中採取不同的覓食策略。例如,某些樹蛙物種能夠在樹冠層中靈活移動,捕食樹葉和樹枝上的昆蟲;而一些地棲蛙類則主要在地面或低矮植被中尋找獵物。植被的結構複雜性對蛙類的覓食效率有重要影響,豐富的植被不僅提供了更多的獵物,還為蛙類提供了隱蔽和等待獵物的場所。
陸地棲息地對蛙類的生存至關重要,因為它提供了躲避天敵的場所。蛙類面臨來自多種捕食者的威脅,包括蛇類、鳥類和哺乳動物。為了應對這些威脅,蛙類進化出了各種適應策略。例如,許多物種具有保護性的體色,能夠與周圍環境融為一體。樹蛙通常具有綠色或棕色的體色,可以很好地隱藏在樹葉或樹皮上。地棲蛙類則可能具有斑駁的體色,以便在落葉或碎石中隱藏。此外,一些蛙類物種會利用地面的裂縫、倒木下的空隙或岩石縫隙作為隱蔽場所。
蛙類作為變溫動物,其體溫受環境溫度的強烈影響。陸地棲息地為蛙類提供了調節體溫的機會。在寒冷季節,蛙類可能會尋找地下洞穴、落葉層或腐爛的木頭等微棲息地來避寒。這些場所不僅提供了相對穩定的溫度,還能保持一定的濕度,防止蛙類脫水。相反,在炎熱的天氣裡,蛙類可能會尋找陰涼潮濕的場所,如岩石下方或濃密植被的遮蔽處。某些物種甚至能夠通過改變體色來調節吸收或反射陽光的程度,從而調節體溫。
季節性遷移是許多蛙類物種利用陸地棲息地的重要方式。在繁殖季節,成年蛙類會從陸地棲息地遷移到水體進行繁殖。這種遷移可能涉及相當長的距離,蛙類需要穿越各種陸地環境。例如,某些蛙類物種可能需要穿越森林、草地甚至是人類居住區才能到達繁殖水體。這種遷移過程中,適宜的陸地棲息地不僅提供了移動通道,還為蛙類提供了短暫的休息和覓食場所。
陸地棲息地的濕度對蛙類至關重要。大多數蛙類物種通過皮膚進行呼吸和水分交換,因此容易脫水。適宜的陸地棲息地應該能夠維持一定的濕度。落葉層、苔蘚覆蓋的地面或濕潤的土壤都是理想的微棲息地。某些適應乾旱環境的蛙類物種進化出了特殊的行為或生理機制來應對低濕度環境,如分泌防水的蠟質物質或在乾季時進入休眠狀態。
人類活動對蛙類的陸地棲息地利用產生了深遠影響。棲息地破壞和破碎化是最直接的威脅。城市化進程導致大片自然棲息地被轉換為建築用地,剩餘的棲息地被道路和其他基礎設施分割。這不僅減少了可用的棲息空間,還阻礙了蛙類的遷移和擴散。農業活動,特別是集約化農業,通過清除自然植被、使用農藥和化肥,也嚴重影響了蛙類的陸地棲息地。
氣候變化正在改變蛙類陸地棲息地的適宜性。溫度升高和降水模式的改變可能導致某些地區變得過於乾燥或過熱,不再適合蛙類生存。這可能迫使某些物種向更適宜的區域遷移,或者面臨局部滅絕的風險。
為了應對這些挑戰,保護和恢復蛙類的陸地棲息地成為保育工作的重點。這包括保護現有的自然棲息地、恢復退化的環境,以及在人為改變的景觀中創造適宜的微棲息地。例如,在城市環境中,通過創建綠色空間、保留天然植被和設置人工濕地,可以為蛙類提供重要的陸地棲息地。在農業區域,鼓勵採用環境友好的耕作方式,如保留田邊樹叢和減少農藥使用,可以顯著改善蛙類的生存環境。
理解蛙類對陸地棲息地的利用模式對於制定有效的保護策略至關重要。這要求我們全面考慮蛙類的生態需求,包括食物來源、避難所、遷移路線和微氣候條件等。只有充分了解蛙類與其陸地棲息地之間的複雜相互作用,我們才能確保這些獨特生物及其所依賴的生態系統的長期生存。
11.3 蛙類微棲息地選擇
蛙類的微棲息地選擇是一個複雜而精細的過程,反映了這些兩棲動物對環境的精確適應。微棲息地是指在較大棲息地範圍內的小尺度環境,通常包括特定的物理結構、微氣候條件和生物因素。蛙類對微棲息地的選擇直接影響其生存、繁殖成功和種群動態。
在水生環境中,蛙類的微棲息地選擇表現得尤為明顯。不同物種的蝌蚪會選擇不同的水體微環境。例如,有些物種的蝌蚪偏好水體表層,這裡光照充足,有利於藻類生長,為蝌蚪提供豐富的食物來源。而其他物種的蝌蚪則可能選擇水體底部,這裡有機碎屑豐富,且捕食風險較低。水生植物的密度和分布也是蝌蚪選擇微棲息地的重要因素。密集的水生植物不僅提供了遮蔽,還能調節局部水質,如增加溶氧量和降低水溫波動。
成年蛙類在水生環境中的微棲息地選擇更為多樣。例如,某些樹蛙物種可能選擇水面上方的植物枝葉作為鳴叫站,這樣既可以有效傳播聲音,又能避免水生捕食者。水蛙屬的一些成員則可能選擇水邊的淺水區域,這裡既可以快速進入水中躲避天敵,又能方便地捕捉水陸兩棲的獵物。
在陸地環境中,蛙類的微棲息地選擇涉及多個方面。地表結構是一個關鍵因素。許多地棲蛙類偏好有落葉層覆蓋的地面,這種環境不僅保持了適宜的濕度,還提供了豐富的食物來源和隱蔽場所。一些蛙類物種則可能選擇岩石縫隙或倒木下的空隙作為日間躲藏處,這些地方通常溫度較低且濕度較高,有助於蛙類避免脫水。
植被結構對蛙類的微棲息地選擇有重要影響。樹蛙科的許多成員展現出對不同高度和類型植被的偏好。有些物種可能選擇樹幹上的特定高度,這裡可能提供了最佳的溫度和濕度組合。其他物種則可能偏好特定的葉片類型或大小,這些可能與其體色或體型相匹配,有利於偽裝。地面層的植被密度和組成也影響著地棲蛙類的微棲息地選擇,密集的地表植被可以為蛙類提供更多的遮蔽和獵物。
微氣候條件是蛙類選擇微棲息地的另一個重要考慮因素。由於蛙類是變溫動物,它們需要通過選擇適當的微棲息地來調節體溫。在寒冷時期,蛙類可能選擇相對溫暖的微環境,如太陽直射的岩石表面或腐爛的木頭內部。在炎熱時期,它們則可能尋找陰涼潮濕的地方,如大樹底部或濃密灌木叢中。濕度是另一個關鍵因素,大多數蛙類需要保持皮膚濕潤以進行呼吸和水分交換。因此,它們通常選擇能維持高濕度的微棲息地,如苔蘚覆蓋的地面或濕潤的洞穴。
食物可用性也影響蛙類的微棲息地選擇。不同的微棲息地可能提供不同類型和數量的獵物。例如,某些蛙類可能選擇靠近水體的陸地區域,這裡通常有大量的飛蟲。其他物種則可能偏好落葉豐富的地面,那裡有豐富的地棲無脊椎動物。
捕食壓力是影響蛙類微棲息地選擇的另一個重要因素。蛙類需要在獲取資源和避免被捕食之間取得平衡。例如,開闊地帶可能提供更多的獵物,但同時也增加了被捕食的風險。因此,許多蛙類物種選擇能夠提供充分遮蔽同時又不過分限制活動的微棲息地。
人類活動對蛙類的微棲息地選擇產生了深遠影響。棲息地破壞和改變可能導致適宜的微棲息地減少或消失。例如,森林砍伐不僅減少了總體棲息地面積,還改變了微氣候條件,如地表溫度和濕度。城市化過程中,不透水表面的增加和自然植被的清除大大減少了適合蛙類的微棲息地。
氣候變化也正在改變微棲息地的適宜性。全球變暖可能導致某些微棲息地變得過熱或過於乾燥。這可能迫使蛙類改變其微棲息地選擇模式,或者面臨無法找到適宜微棲息地的風險。
理解蛙類的微棲息地選擇對於制定有效的保護策略至關重要。這要求我們不僅要保護大尺度的棲息地,還要關注維護和恢復適宜的微棲息地。例如,在城市綠地設計中,可以通過創造多樣化的微環境,如保留落葉、設置小型水體和維護不同層次的植被,來滿足不同蛙類物種的需求。在農業景觀中,保留田邊樹叢、石堆或濕地斑塊可以為蛙類提供重要的微棲息地。
通過深入研究蛙類的微棲息地選擇,我們不僅能更好地理解這些生物的生態需求,還能為整個生態系統的管理和恢復提供重要見解。只有充分考慮到微棲息地的重要性,我們才能確保蛙類及其所依賴的生態系統的長期健康和穩定。
11.4 都市化對蛙類棲息地的影響
都市化對蛙類棲息地的影響是一個複雜而深遠的問題,涉及多個生態和環境因素的改變。隨著城市的擴張和發展,自然棲息地被轉變為人工環境,這對依賴特定生態條件的蛙類造成了巨大影響。
首先,都市化最直接的影響是棲息地的喪失和破碎化。大規模的建設活動導致濕地、森林和草地等自然環境被清除,取而代之的是建築物、道路和其他人工結構。這不僅減少了蛙類可用的棲息空間,還將原本連續的棲息地分割成孤立的小斑塊。例如,一個原本完整的濕地系統可能被道路和建築分割成幾個小型水體,這些孤立的水體可能無法支持健康的蛙類種群。棲息地的破碎化還阻礙了蛙類的遷移和擴散,增加了局部種群滅絕的風險。
水文條件的改變是都市化對蛙類棲息地影響的另一個重要方面。城市發展通常伴隨著大量不透水表面的增加,如瀝青路面和混凝土建築。這些表面阻礙了雨水滲入地下,改變了自然的水文循環。結果是,一方面降雨迅速形成地表徑流,可能導致短暫的洪水;另一方面,乾旱期間的地下水補給減少,可能導致小型水體乾涸。對蛙類而言,這種水文條件的不穩定性可能嚴重影響其繁殖成功率,特別是那些依賴臨時性水體繁殖的物種。
水質污染是都市化環境中蛙類面臨的另一大挑戰。城市地表徑流常常攜帶各種污染物,包括重金屬、石油產品、殺蟲劑和肥料等,這些物質最終進入水體,危害蛙類及其食物來源。某些污染物,如內分泌干擾物,可能直接影響蛙類的生理功能和繁殖能力。此外,城市水體的富營養化問題也很普遍,這可能導致有害藻類的大量繁殖,降低水體的溶氧量,進而影響蛙類和其他水生生物的生存。
微氣候的改變是都市化對蛙類棲息地影響的一個較為隱蔽但同樣重要的方面。城市熱島效應導致城市地區的溫度通常高於周圍鄉村地區。對於蛙類這樣的變溫動物來說,這種溫度升高可能超出其生理耐受範圍,特別是在夏季。同時,城市環境通常比自然環境乾燥,這對需要保持皮膚濕潤的蛙類來說是一個挑戰。微氣候的改變可能迫使蛙類改變其活動模式或尋找新的棲息地。
光污染和噪音污染是都市化環境中的另外兩個問題。夜間的人工照明可能干擾蛙類的自然行為模式,如覓食和繁殖活動。某些蛙類物種依賴月光或星光來同步其繁殖活動,人工照明可能干擾這一過程。噪音污染,特別是來自交通和建築活動的噪音,可能干擾蛙類的聲學通訊。這對依賴鳴叫來吸引配偶的物種尤其不利。
都市化還帶來了外來物種入侵的風險增加。城市環境常常成為外來物種的引入和擴散中心。這些外來物種可能與本地蛙類競爭資源,捕食本地蛙類,或引入新的疾病。例如,在許多城市水體中,外來的牛蛙(Rana catesbeiana)成為優勢物種,對本地蛙類造成嚴重威脅。
然而,都市化對蛙類的影響並非完全負面。某些適應能力強的蛙類物種能夠在城市環境中生存甚至繁榮。城市公園、花園和人工水體可能為這些物種提供新的棲息地。例如,某些樹蛙物種能夠很好地適應城市環境,利用人工水體進行繁殖。這種適應性提示我們,通過適當的城市規劃和管理,可以在一定程度上減輕都市化對蛙類的負面影響。
為了減輕都市化對蛙類棲息地的影響,一些城市已經開始採取措施。例如,建立城市濕地公園,不僅為蛙類提供棲息地,還能改善城市水質和微氣候。綠色基礎設施的建設,如雨水花園和生物滯留池,可以幫助管理城市地表徑流,同時為蛙類創造適宜的微棲息地。在新區開發中保留自然水體和綠地,並確保它們之間的連通性,也是重要的保護策略。
此外,提高公眾對城市生物多樣性的認識和參與也很重要。鼓勵居民在自家花園中創造適合蛙類的環境,如小型池塘或濕地區域,可以在社區層面上為蛙類提供棲息地網絡。公民科學項目,如城市蛙類監測計劃,不僅能收集寶貴的生態數據,還能增強公眾對蛙類保護的意識和參與度。
理解並減輕都市化對蛙類棲息地的影響,需要城市規劃者、生態學家和公眾的共同努力。通過綜合考慮蛙類的生態需求和城市發展的需要,我們可以創造更加生態友好的城市環境,不僅有利於蛙類的保護,還能提高城市的生態韌性和宜居性。
11.5 打造蛙類友善環境
打造蛙類友善環境是一項重要的生態保育工作,不僅有助於維護生物多樣性,還能為人類創造更加和諧的生活空間。在都市化快速發展的今天,許多蛙類的棲息地受到嚴重威脅,因此我們需要採取積極行動來創造適合蛙類生存的環境。
首先,水源的保護和管理是打造蛙類友善環境的關鍵。蛙類的生活周期與水密不可分,從產卵到蝌蚪階段都需要水環境。因此,保護和恢復自然水體,如小溪、池塘和濕地,對蛙類至關重要。在都市地區,我們可以設計和建造人工濕地或雨水花園,這些設施不僅能為蛙類提供棲息地,還能幫助淨化水質和調節城市微氣候。
其次,維護和創造多樣化的陸地棲息環境同樣重要。成年蛙類需要適當的陸地環境來覓食、躲避天敵和度過非繁殖季節。我們可以在公園、社區綠地和私人花園中種植本地植物,創造層次豐富的植被結構。落葉、枯木和石塊堆等自然元素也應該保留,因為它們為蛙類提供了重要的庇護所和覓食場所。
減少化學品的使用是打造蛙類友善環境的另一個重要方面。蛙類的皮膚非常敏感,容易受到農藥、除草劑和其他化學污染物的傷害。在園藝和農業實踐中,我們應該盡量採用有機方法,避免使用對蛙類有害的化學品。此外,適當管理雨水徑流,防止污染物進入水體,也是保護蛙類棲息地的重要措施。
光污染對蛙類的生活也有顯著影響。許多蛙類是夜行性動物,過度的人工照明會干擾它們的活動模式和繁殖行為。在設計戶外照明時,我們應該考慮使用定向燈具,避免光線溢出,並在非必要時段減少照明強度或完全關閉燈光。
此外,建立生態廊道對於打造蛙類友善環境也很重要。由於城市化導致棲息地破碎化,蛙類種群之間的基因交流受到限制。通過設計和實施生態廊道,如綠色通道或生態橋樑,我們可以幫助蛙類在不同棲息地之間安全移動,促進種群交流和基因多樣性。
教育和公眾參與是成功打造蛙類友善環境的關鍵因素。通過舉辦社區活動、自然教育課程和公民科學項目,我們可以提高公眾對蛙類保護的認知和參與度。例如,組織夜間觀蛙活動,讓人們親身體驗蛙類的魅力;或者發起社區蛙類棲息地修復項目,讓居民直接參與到環境改善工作中。
在城市規劃和建築設計中融入蛙類友善理念也很重要。例如,在新建住宅區或商業區中規劃生態池塘和綠色空間,或者在建築物外牆和屋頂設計垂直綠化系統,這些都可以為蛙類創造棲息環境。同時,在道路設計中考慮設置蛙類通道,減少道路對蛙類遷移的阻隔效應。
最後,持續監測和適應性管理是維護蛙類友善環境的重要手段。通過定期調查蛙類種群數量和分佈情況,我們可以及時發現問題並作出調整。例如,如果發現某些措施效果不佳,可以嘗試新的方法;如果某些地區的蛙類數量明顯減少,則需要深入研究原因並採取針對性的保護措施。
打造蛙類友善環境是一項長期的工作,需要政府、科研機構、社區組織和公眾的共同努力。通過綜合考慮水源保護、棲息地多樣化、化學品管理、光污染控制、生態廊道建設、教育宣傳、城市規劃和持續監測等多個方面,我們可以為蛙類創造更加適宜的生存環境,同時也為人類塑造一個更加生態、宜居的生活空間。
11.6 營造蛙類棲息地
人為營造蛙類棲息地以吸引野生蛙類到生態觀察園是一項富有挑戰性但極具意義的工作。這不僅能為蛙類提供安全的棲息環境,還能為生態教育和科研觀察提供寶貴的場所。要成功實現這一目標,需要綜合考慮多個因素,並採取一系列精心設計的措施。
首先,水體的設計和管理是吸引蛙類的關鍵。在生態觀察園中,應該創造多樣化的水環境,包括淺水區、深水區、緩流區和靜水區。這種多樣性能滿足不同蛙類的需求,例如樹蛙可能偏好淺水區域,而青蛙則可能更喜歡深水區。水體的邊緣應該呈不規則形狀,並設置緩坡,便於蛙類進出水體。此外,保持水質清潔至關重要,可以通過種植水生植物如睡蓮、菖蒲等來淨化水質,同時這些植物還能為蛙類提供隱蔽和產卵場所。
陸地環境的營造同樣重要。在水體周圍應該種植多層次的植被,包括地被植物、灌木和喬木。這種複雜的植被結構不僅能為蛙類提供庇護所,還能吸引昆蟲,為蛙類提供食物來源。選擇植物時應優先考慮本地物種,因為它們更適應當地環境,也更有利於維持生態平衡。例如,在台灣的生態觀察園中,可以種植山黃麻、楝樹等本地植物。
在設計棲息地時,要考慮到不同蛙類的特殊需求。例如,樹蛙需要垂直的結構來攀爬,可以在園區內設置一些直立的枯木或者特製的攀爬架。對於喜歡躲藏的蛙類,可以在地面上放置一些石塊、樹皮或朽木,為它們提供隱蔽處。這些結構不僅能滿足蛙類的需求,還能增加園區的自然美感。
食物供應是吸引和留住蛙類的另一個重要因素。大多數蛙類以昆蟲為食,因此可以通過一些方法來吸引昆蟲。例如,種植一些能吸引蝴蝶和蜜蜂的花卉,或者在園區內設置一些腐爛的水果,這些都能吸引各種昆蟲,進而吸引蛙類。但要注意控制昆蟲的數量,避免對周圍環境造成負面影響。
光照管理對於吸引蛙類也很重要。許多蛙類是夜行性動物,過強的人工照明可能會干擾它們的活動。因此,在設計園區照明時,應該採用低強度、黃色的燈光,並將燈具安裝在較低的位置,避免直接照射水面和主要活動區域。在不影響安全的情況下,可以考慮在夜間完全關閉部分區域的照明。
聲音環境的營造也不容忽視。蛙類通過鳴叫來進行交流和吸引配偶,因此一個安靜的環境對它們來說很重要。可以通過種植密集的植被來減少外界噪音的干擾,或者在園區周圍設置隔音屏障。同時,可以考慮播放錄製的蛙鳴聲音,這可能會吸引野生蛙類前來查看。
為了確保蛙類的安全,需要採取一些保護措施。例如,在園區周圍設置柵欄,防止捕食者如蛇、貓等進入。但柵欄的設計要合理,不能阻礙小型動物的自由進出。此外,嚴格控制園區內的農藥和化學品使用,採用有機的園藝管理方法,以保護蛙類脆弱的皮膚。
在營造棲息地的過程中,模仿自然環境的季節性變化也很重要。例如,可以通過調節水位來模擬雨季和乾季的變化,這對於刺激某些蛙類的繁殖行為可能很有幫助。同時,保留一些落葉和枯枝,不要過度整理環境,這樣更接近蛙類在自然界中的生存環境。
為了吸引多樣化的蛙類,可以在園區內創造微棲息地。例如,為樹蛙設置高大的植物或人工結構,為喜歡潮濕環境的蛙類創造苔蘚覆蓋的石頭區域,為喜歡曬太陽的蛙類提供開闊的岩石或木頭平台。這種多樣化的環境設計能夠滿足不同蛙類的需求,增加整個生態系統的豐富度。
在生態觀察園中設置觀察設施時,需要特別小心,以免干擾蛙類的自然行為。可以設計隱蔽的觀察點,如使用單向玻璃的觀察亭或高架步道,讓遊客能夠近距離觀察蛙類而不會驚擾它們。同時,可以安裝一些水下攝像機或夜視攝像機,讓遊客能夠觀察到蛙類的水下活動和夜間行為。
最後,持續的監測和管理是確保生態觀察園成功的關鍵。定期進行蛙類種群調查,記錄出現的物種和數量變化。根據監測結果,及時調整管理策略,例如改變水體管理方式、調整植被結構或增添新的微棲息地。同時,與當地的生態學家和保育專家保持密切合作,獲取專業建議和最新的研究成果,不斷完善園區的管理方法。
通過這些精心設計和管理的措施,生態觀察園能夠成功吸引多樣化的野生蛙類,為它們提供安全的棲息環境,同時也為公眾提供一個親近自然、學習生態知識的絕佳場所。這不僅有助於蛙類保護,還能提高公眾的環境意識,促進人與自然的和諧共處。
12. 蛙類威脅與保育
蛙類作為生態系統中的重要成員,近年來面臨著嚴峻的生存威脅。棲息地喪失和破碎化是最主要的問題之一,隨著城市化進程加快和農業用地擴張,許多蛙類賴以生存的濕地和森林遭到破壞。污染也對蛙類造成巨大傷害,農藥、工業廢水和城市排放物不僅直接毒害蛙類,還破壞了它們的生存環境。
氣候變遷帶來的影響同樣不容忽視,極端天氣事件的增加和溫度變化影響了蛙類的繁殖周期和分布範圍。此外,全球範圍內肆虐的兩棲類疾病,如由壺菌引起的壺菌病,已經導致多個物種的滅絕或瀕危。
面對這些威脅,科學家和保育人士正在採取多方面的行動。首先是加強對現有棲息地的保護,建立更多的自然保護區和生態廊道。其次,開展了大規模的棲息地修復計劃,如濕地重建和森林恢復。在疾病防控方面,研究人員正在努力開發疫苗和治療方法,同時加強對野生種群的健康監測。
公眾教育和參與也是蛙類保育的重要組成部分。通過環境教育項目和公民科學計劃,提高人們對蛙類保護的認識和參與度。此外,國際合作在應對跨境威脅方面發揮著關鍵作用,如控制非法野生動物貿易和應對全球性疾病傳播。
儘管面臨諸多挑戰,但一些保育努力已經取得了顯著成效。例如,在澳大利亞,通過建立專門的繁殖設施和棲息地保護,成功挽救了瀕臨滅絕的柯氏樹蛙。這些成功案例為蛙類的未來保育工作提供了希望和經驗。
12.1 蛙類棲息地喪失與破碎化
蛙類棲息地喪失與破碎化是當前威脅蛙類生存的最嚴重問題之一,這一現象在全球範圍內普遍存在,並對蛙類種群造成了深遠的影響。棲息地喪失指的是蛙類原有的生存環境被完全破壞或改變,而棲息地破碎化則是指原本連續的大面積棲息地被分割成多個小區域,彼此之間相互隔離。這兩種情況往往同時發生,共同對蛙類的生存構成嚴重威脅。
造成蛙類棲息地喪失與破碎化的主要原因是人類活動的擴張。城市化進程的加快導致大量農田、森林和濕地被轉化為建築用地。農業的集約化和擴張也使得許多自然棲息地被改造成單一作物的種植區。此外,道路、鐵路等基礎設施的修建不僅直接佔用了蛙類的棲息地,還將原本完整的生態系統切割成碎片。在一些地區,濫伐森林、過度放牧和礦產開發等活動也嚴重破壞了蛙類的生存環境。
棲息地喪失對蛙類的影響是直接而嚴重的。當一片濕地被填平或一片森林被砍伐時,生活在其中的蛙類種群可能會在短時間內完全消失。例如,在巴西亞馬遜地區,大規模的森林砍伐導致許多特有的樹蛙物種數量急劇下降,有些甚至面臨滅絕的威脅。棲息地的喪失不僅直接減少了蛙類的生存空間,還破壞了它們賴以生存的食物網和生態系統平衡。
棲息地破碎化的影響則更加複雜和長期。當一片大的棲息地被分割成多個小區域時,每個小區域能夠支持的蛙類種群規模就會減小。這種規模的縮小會增加種群滅絕的風險,特別是在面臨疾病、氣候變化或其他環境壓力時。此外,棲息地之間的隔離會阻礙蛙類的遷移和基因交流,導致近親繁殖和遺傳多樣性的降低,從而降低種群的適應能力和長期生存機會。
在台灣,隨著經濟的快速發展,蛙類棲息地喪失與破碎化的問題也日益嚴重。許多低地的濕地和水田被改造成工業區或住宅區,山區的森林也因道路建設和觀光開發而遭到分割。這對台灣的特有蛙類,如梭德氏赤蛙和莫氏樹蛙等,造成了嚴重的威脅。台北赤蛙就是一個典型的例子,由於都市化導致的棲息地喪失,這種曾經在台北盆地普遍存在的蛙類現在已經極為罕見。
棲息地喪失與破碎化還會導致蛙類的遷移行為受阻。許多蛙類需要在不同的棲息地之間遷移以完成其生活史,例如從繁殖地到覓食地或冬眠地的季節性遷移。當這些棲息地之間的連接被切斷時,蛙類的正常生活周期就會受到干擾。在一些地區,每年都有大量蛙類在穿越道路時被車輛碾壓,這種現象正是棲息地破碎化的直接後果。
氣候變化加劇了棲息地喪失與破碎化對蛙類的影響。隨著全球氣溫升高,許多蛙類需要向高緯度或高海拔地區遷移以尋找適宜的環境。然而,如果棲息地已經高度破碎化,這種遷移就變得極其困難,可能導致一些物種無法適應氣候變化而滅絕。
為了應對蛙類棲息地喪失與破碎化的問題,科學家和保育人士採取了多種策略。首先是加強對現有棲息地的保護,建立更多的自然保護區和生態廊道。例如,在台灣的國家公園和自然保留區內,對蛙類棲息地的保護得到了加強。其次是開展棲息地修復和重建工作,如濕地恢復和森林重植。在一些地區,人工建造的生態池塘和濕地成功地為蛙類提供了新的棲息地。
在城市規劃和基礎設施建設中納入生態考量也是減少棲息地破碎化的重要方法。例如,在道路設計中增加生態通道或涵洞,可以幫助蛙類安全穿越道路,減少棲息地隔離的影響。在農業區,鼓勵農民保留田間小水塘和植被帶,為蛙類提供棲息和繁殖場所。
此外,提高公眾意識和參與度對於蛙類棲息地的保護至關重要。通過環境教育和公民科學項目,讓更多人了解蛙類棲息地保護的重要性,並參與到保護行動中來。例如,在台灣,一些社區組織了「護蛙巡邏隊」,在蛙類遷徙季節幫助它們安全穿越道路。
最後,長期監測和研究對於理解和應對棲息地喪失與破碎化的影響也很重要。通過持續追蹤蛙類種群動態和棲息地變化,科學家可以更好地評估保護措施的效果,並及時調整保育策略。
蛙類棲息地喪失與破碎化是一個複雜的問題,需要政府、科研機構、非政府組織和公眾的共同努力才能得到有效解決。雖然挑戰巨大,但通過綜合採取保護、修復、規劃和教育等措施,我們仍然有希望為蛙類創造一個更加安全和連通的生存環境,確保它們能夠繼續在生態系統中發揮重要作用。
12.2 蛙類受疾病的影響
疾病對全球蛙類種群造成的影響是一個日益嚴重的問題,其中尤以壺菌病(Chytridiomycosis)最為突出。這種由壺菌(Batrachochytrium dendrobatidis)引起的真菌感染疾病,自1990年代末被發現以來,已經成為導致全球兩棲類大規模滅絕的主要原因之一。
壺菌病的傳播速度驚人,影響範圍廣泛。這種真菌能夠感染蛙類的皮膚,干擾其正常的滲透調節功能,最終導致感染個體死亡。由於蛙類的皮膚在呼吸和水分平衡中扮演著關鍵角色,壺菌的感染對它們的生理功能造成了毀滅性的打擊。這種疾病已經在全球六大洲的蛙類中被發現,造成了多個物種的滅絕或瀕危。
以澳大利亞的雨蛙(Litoria sp.)為例,自壺菌病爆發以來,多個物種的數量急劇下降,其中一些甚至完全消失在野外。在中美洲,曾經普遍存在的金蛙(Atelopus zeteki)也因壺菌病而瀕臨滅絕,目前只能在人工繁育設施中維持種群。
除了壺菌病,蛙類還面臨著其他疾病的威脅。例如,由類壺菌(Batrachochytrium salamandrivorans)引起的疾病主要影響蠑螈和蜥蜴,但也可能對某些蛙類構成威脅。此外,藍舌病毒(Ranavirus)也是一種常見的兩棲類病原體,能夠導致大規模的死亡事件。
疾病對蛙類的影響不僅限於直接導致死亡,還會削弱種群的整體健康狀況和繁殖能力。受感染的個體可能表現出行為異常,如減少覓食活動或改變繁殖行為,這些都會對種群的長期生存造成不利影響。
氣候變化進一步加劇了疾病對蛙類的威脅。溫度和濕度的變化可能有利於某些病原體的生存和傳播,同時也可能降低蛙類的免疫力。例如,研究表明,在某些地區,氣溫升高與壺菌病爆發之間存在關聯。
人類活動也在疾病傳播中扮演了重要角色。全球化和國際貿易增加了病原體跨區域傳播的風險。例如,用於實驗室研究或寵物貿易的蛙類可能在無意中將疾病帶到新的地區。此外,棲息地破壞和污染等因素也會降低蛙類的抵抗力,使它們更容易受到疾病的侵襲。
面對疾病的威脅,科研人員和保育工作者採取了多種應對策略。首先是加強對野生蛙類種群的健康監測。通過定期採樣和檢測,可以及早發現疾病爆發,並採取相應的防控措施。例如,在澳大利亞,研究人員開發了快速檢測壺菌的方法,大大提高了疾病監測的效率。
其次,科學家們正在積極研發治療方法和預防措施。對於壺菌病,一些研究團隊嘗試使用抗真菌藥物進行治療,或者通過提高環境溫度來抑制真菌的生長。在預防方面,研究人員正在探索疫苗的可能性,雖然目前還沒有廣泛應用的疫苗,但一些初步研究結果顯示了希望。
遷地保育也是應對疾病威脅的重要手段。對於受疾病嚴重威脅的物種,建立人工繁育計劃可以確保它們不會完全滅絕。例如,在巴拿馬,科學家們建立了金蛙的人工繁育設施,以保存這個瀕危物種。這些設施不僅可以維持種群,還可以進行疾病研究和治療實驗。
改善蛙類的棲息環境也是提高其抵抗疾病能力的關鍵。健康的生態系統可以為蛙類提供更好的食物來源和更少的環境壓力,從而增強它們的免疫力。因此,棲息地保護和修復工作對於疾病防控也具有重要意義。
國際合作在應對全球性的疾病威脅中發揮著關鍵作用。科學家們通過跨國合作項目共享研究成果和資源,加速了對疾病的了解和防控措施的開發。例如,「兩棲類疾病全球追蹤系統」(Amphibian Disease Global Tracking System)就是一個國際合作平台,匯集了全球範圍內的兩棲類疾病數據。
公眾教育和參與同樣重要。提高人們對蛙類疾病的認識,可以減少人為傳播疾病的風險。例如,教育遠足者和生態旅遊者在不同區域之間轉移時清潔鞋子和裝備,可以降低將病原體帶入新區域的可能性。
雖然疾病對蛙類構成了嚴重威脅,但科學界的不懈努力也帶來了一些積極的進展。例如,在澳大利亜西部,研究人員發現一些蛙類種群已經開始對壺菌產生抗性。這種自然選擇的過程給蛙類的長期生存帶來了希望。
儘管如此,疾病仍然是蛙類面臨的主要威脅之一,需要持續的關注和研究。只有通過多方面的努力,包括疾病監測、治療研究、棲息地保護、國際合作和公眾教育等,我們才能有效地減輕疾病對蛙類的影響,確保這些重要的生態系統成員能夠繼續在地球上繁衍生息。
12.3 氣候變遷對蛙類的影響
氣候變遷對蛙類的影響是多方面且深遠的,這種全球性的環境變化正在以多種方式威脅著蛙類的生存和繁衍。蛙類作為變溫動物,其生理功能和行為模式與環境溫度密切相關,因此對氣候變化特別敏感。
首先,全球氣溫上升直接影響了蛙類的分布範圍。許多蛙類物種正在向高緯度或高海拔地區遷移,以尋找適合的溫度環境。例如,在哥斯大黎加的蒙特維德雲霧森林中,研究人員發現一些低海拔的蛙類物種正逐漸向更高海拔遷移。然而,並非所有蛙類都能成功適應這種遷移。對於那些已經生活在山頂或極地地區的蛙類來說,他們可能面臨「無處可去」的困境,最終導致種群數量的下降甚至滅絕。
氣溫升高還會影響蛙類的繁殖週期和成功率。許多蛙類的繁殖行為是由溫度變化觸發的,氣候變遷可能導致繁殖時間的提前或延後,使其與食物來源或其他重要生態因素不同步。例如,在英國,研究人員發現普通蟾蜍(Bufo bufo)的繁殖季節比40年前提前了約10天。如果這種變化導致蝌蚪孵化時間與其主要食物來源藻類的生長高峰不一致,可能會嚴重影響蝌蚪的存活率。
此外,氣溫升高還可能影響蛙卵的發育。許多蛙類的卵需要特定的溫度範圍才能正常發育,溫度過高可能導致卵的死亡或畸形。例如,在澳大利亞,研究人員發現當溫度超過某個閾值時,條紋沼蛙(Limnodynastes peronii)的卵會出現更高的畸形率。
降水模式的改變是氣候變遷對蛙類影響的另一個重要方面。許多地區正經歷著降水量的顯著變化,有些地方降水增加,而有些地方則面臨乾旱。這些變化直接影響了蛙類的生存環境。例如,在乾旱加劇的地區,許多臨時性水體可能會完全乾涸,導致依賴這些水體繁殖的蛙類失去繁殖場所。反之,在降水增加的地區,過多的降雨可能會沖走蛙卵或改變水體的化學成分,同樣影響蛙類的繁殖成功率。
極端天氣事件的增加也給蛙類帶來了嚴峻的挑戰。頻繁的熱浪、洪水或寒潮可能直接導致大量蛙類死亡。例如,在2019年澳大利亞的極端高溫天氣中,研究人員發現許多蛙類因無法承受高溫而死亡。這些極端事件不僅直接威脅蛙類的生存,還可能破壞其棲息地,進一步加劇種群的脆弱性。
氣候變遷還間接影響了蛙類的食物網。溫度和降水的變化可能改變昆蟲和其他無脊椎動物的豐度和分布,這些生物是許多蛙類的主要食物來源。同時,氣候變化也可能影響捕食蛙類的動物,改變整個生態系統的平衡。
值得注意的是,氣候變遷還可能加劇疾病對蛙類的影響。例如,研究表明,氣候變暖可能有利於某些病原體的生存和傳播,如壺菌(Batrachochytrium dendrobatidis)。同時,氣候壓力可能降低蛙類的免疫力,使其更容易感染疾病。
面對氣候變遷的威脅,科學家們正在採取多種策略來幫助蛙類適應和生存。其中一個重要方向是識別和保護氣候避難所。這些地區通常具有相對穩定的微氣候,可以為蛙類提供庇護。例如,在澳大利亞的熱帶雨林中,研究人員發現一些深谷和高海拔地區可能成為蛙類的氣候避難所,並建議優先保護這些區域。
另一個策略是通過棲息地管理來減輕氣候變遷的影響。例如,在一些地區,科學家們嘗試通過種植適當的植被來為蛙類創造更多的遮蔽和降溫區域。在乾旱地區,人工維護或創造水體也是一種重要的保護措施。
基因保護也是應對氣候變遷的一個重要方向。通過保存不同種群的遺傳多樣性,可以增加蛙類對環境變化的適應能力。一些研究項目正在收集和保存瀕危蛙類的基因材料,以備將來可能的重新引入或基因強化。
監測和預測也是應對氣候變遷影響的關鍵。通過長期監測蛙類種群和其棲息地的變化,科學家們可以更好地理解氣候變遷的影響,並制定相應的保護策略。同時,利用氣候模型預測未來的變化趨勢,可以幫助我們提前採取保護措施。
此外,減少溫室氣體排放、保護和恢復自然生態系統等全球性的氣候行動,對於減緩氣候變遷對蛙類的影響至關重要。這需要政府、企業和公眾的共同努力。
儘管氣候變遷對蛙類構成了嚴峻的挑戰,但一些蛙類物種也表現出了適應能力。例如,在澳大利亞,研究人員發現一些蛙類物種能夠通過改變其行為模式(如改變活動時間)或生理特徵(如增強耐熱能力)來適應氣候變化。這些發現為蛙類的未來生存提供了一絲希望。
然而,我們必須認識到,並非所有蛙類物種都能成功適應如此快速的環境變化。因此,全面而持續的保護行動仍然是確保蛙類在氣候變遷下生存的關鍵。這需要科研人員、保育工作者、政策制定者和公眾的共同努力,以確保這些重要的生態指標物種能夠繼續在我們的生態系統中扮演重要角色。
12.4 環境污染對蛙類的影響
環境污染對蛙類的影響是多方面且深遠的,這種人為造成的生態威脅正在全球範圍內嚴重危及蛙類的生存。蛙類由於其特殊的生理特徵和生活習性,對環境污染尤為敏感,常被視為生態系統健康狀況的指標生物。
水污染是影響蛙類最直接且最嚴重的污染形式之一。蛙類的生活周期大部分依賴於水環境,從產卵、蝌蚪發育到成年個體的生存都與水密不可分。工業廢水、農業排放物和城市污水中的有毒物質可以直接毒害蛙類,導致死亡或畸形。例如,在美國加利福尼亞州的農業地區,研究人員發現由於農藥污染,當地的紅腿蛙(Rana aurora)種群數量急劇下降,許多個體出現畸形。這些污染物不僅影響成年蛙類,對蝌蚪的影響更為顯著。水體中的重金屬、農藥殘留和內分泌干擾物質會干擾蝌蚪的正常發育,導致生長遲緩、免疫力下降,甚至引發基因突變。
農藥污染是另一個嚴重威脅蛙類的環境問題。大量使用的殺蟲劑、除草劑和殺菌劑不僅直接毒害蛙類,還會通過食物鏈富集,最終對蛙類造成更大的傷害。一些研究表明,即使是低濃度的農藥也可能影響蛙類的內分泌系統,干擾其生殖和發育過程。例如,廣泛使用的除草劑草甘膦被發現可以干擾蛙類的性激素水平,影響其性別分化和繁殖能力。在阿根廷的一項研究中,科學家發現暴露於草甘膦的蝌蚪更容易發生畸形,並且存活率顯著降低。
大氣污染也對蛙類產生了重要影響。酸雨是一個典型的例子,它可以直接改變水體和土壤的pH值,影響蛙類的生存環境。酸化的水體會干擾蛙卵的正常發育,降低孵化率,同時也會影響水生植物和微生物的生長,間接影響蛙類的食物來源。此外,大氣中的其他污染物,如氮氧化物和硫氧化物,也可能通過沉降進入水體,對蛙類造成傷害。
塑料污染是近年來越來越受到關注的環境問題,它同樣對蛙類構成威脅。微塑料可以被蛙類誤食,堆積在消化系統中,影響其正常攝食和消化。更為嚴重的是,塑料中的添加劑和吸附的其他污染物可能會干擾蛙類的內分泌系統。在英國的一項研究中,科學家在野外採集的蛙類樣本中檢測到了微塑料,這表明塑料污染已經開始影響野生蛙類種群。
光污染是一種常被忽視但對蛙類影響顯著的污染形式。許多蛙類是夜行性動物,它們的繁殖行為和覓食活動往往在夜間進行。過度的人工照明可能會干擾蛙類的自然行為模式,影響它們的繁殖成功率。例如,在城市化地區,強烈的人工照明可能會吸引蛙類遠離其自然棲息地,增加它們被捕食或遭遇交通事故的風險。
噪音污染也對蛙類產生了負面影響。蛙類依賴聲音進行交流和尋找配偶,環境中的噪音干擾可能會掩蓋蛙類的叫聲,影響它們的繁殖行為。在巴西的一項研究中,科學家發現靠近公路的蛙類種群的叫聲頻率和持續時間都發生了變化,這可能是為了適應交通噪音而進行的調整。
值得注意的是,不同類型的污染往往是相互作用的,它們的綜合影響可能比單一污染源更為嚴重。例如,氣候變化可能會加劇某些污染物的毒性,而棲息地破壞則可能減少蛙類躲避污染的安全區域。
面對環境污染的威脅,科學家和環保工作者正在採取多種措施來保護蛙類。首先是加強環境監測和污染控制。通過建立完善的水質監測系統,及時發現和處理水污染問題。同時,推廣環保型農業實踐,減少農藥和化肥的使用,可以有效降低農業污染對蛙類的影響。
其次,開展生態修復工作也是減輕污染影響的重要手段。例如,在一些受污染嚴重的地區,科學家們嘗試通過種植特定植物來吸收和降解污染物,為蛙類創造更加潔淨的生存環境。在美國的一些礦區,研究人員成功地利用濕地植物來清除水體中的重金屬污染,為當地蛙類種群的恢復創造了條件。
提高公眾意識和參與度也是應對環境污染的關鍵。通過環境教育項目,讓更多人了解環境污染對蛙類和整個生態系統的影響,從而促使人們在日常生活中採取更環保的行為。例如,減少塑料使用、正確處理廢棄物、選擇環保產品等。
立法和政策支持同樣重要。通過制定和執行嚴格的環境保護法規,限制有害物質的排放,可以從源頭上減少污染。例如,歐盟的化學品註冊、評估、授權和限制法規(REACH)就對許多可能危害環境的化學物質的使用進行了嚴格管控。
此外,開展針對性的研究也是應對污染威脅的重要手段。通過深入研究不同污染物對蛙類的影響機制,科學家們可以開發出更有效的保護策略。例如,一些研究團隊正在探索使用生物標記物來早期檢測污染對蛙類的影響,這可以幫助我們更快地發現和應對環境問題。
雖然環境污染對蛙類構成了嚴重威脅,但通過科學研究、政策支持和公眾參與,我們仍有希望減輕這些影響,為蛙類創造一個更加清潔、安全的生存環境。保護蛙類不僅關乎這些生物本身的福祉,也是維護整個生態系統健康的重要一環。
12.5 蛙類保育成功案例
在全球範圍內,儘管蛙類面臨著諸多威脅,但通過科學家、保育人士和公眾的共同努力,已經取得了一些令人鼓舞的保育成功案例。這些案例不僅展示了人類在保護生物多樣性方面的決心和能力,也為其他瀕危物種的保育工作提供了寶貴的經驗和希望。
一個著名的成功案例是澳大利亞的柯氏樹蛙(Litoria cooloolensis)的拯救。這種樹蛙原本僅分布在昆士蘭州東南部的一小片區域,由於棲息地破壞和疾病的影響,其野外種群在20世紀90年代末幾乎滅絕。面對這種情況,澳大利亞的科學家們迅速行動,開展了一項綜合性的保育計劃。他們首先在野外收集了剩餘的個體,建立了人工繁育設施。通過精心的飼養和繁殖管理,科學家們成功地增加了柯氏樹蛙的數量。同時,他們也致力於改善野外棲息地的條件,包括控制入侵物種和恢復原生植被。經過多年的努力,柯氏樹蛙的數量逐漸恢復,並成功地重新引入到野外。這個案例展示了人工繁育結合棲息地保護的策略在挽救瀕危蛙類方面的巨大潛力。
另一個令人振奮的例子來自中美洲的巴拿馬黃金蛙(Atelopus zeteki)。這種鮮艷奪目的蛙類曾經是巴拿馬的國寶,但由於壺菌病的蔓延,其野外種群在2006年後幾乎消失。面對這一危機,保育人士迅速採取行動,在壺菌病到達之前,從野外收集了一批健康的個體。這些蛙被送往美國和巴拿馬的動物園和研究機構,開始了一項大規模的人工繁育計劃。通過精心的照料和管理,黃金蛙在人工環境下成功繁衍。同時,科學家們也在努力研究如何控制壺菌病,並尋找可能的重新引入地點。雖然目前黃金蛙還未能重返野外,但人工種群的建立為這個物種的長期生存提供了希望。
在歐洲,西班牙的中華(Ates muletensis)的保育故事也值得一提。這種蛙類是西班牙馬略卡島的特有種,曾因棲息地喪失和入侵物種的影響而瀕臨滅絕。為了挽救這個物種,西班牙政府和保育組織聯手展開了一系列行動。他們不僅建立了人工繁育計劃,還採取了棲息地恢復和入侵物種控制等措施。特別值得一提的是,他們成功地消除了對中華構成威脅的入侵蛙類,並恢復了一些關鍵的繁殖地。通過這些努力,中華的野外種群數量逐漸回升,成為歐洲兩棲類保育的一個標誌性成功案例。
在北美,加州紅腿蛙(Rana draytonii)的保育故事也展示了長期努力的成果。這種蛙類曾經廣泛分布在加利福尼亞州,但由於棲息地喪失、入侵物種和疾病等因素,其數量急劇下降。保育工作者採取了多管齊下的策略,包括棲息地保護和恢復、入侵物種控制、疾病監測和管理等。特別是,他們在多個地點重建了適合紅腿蛙生存的濕地環境,並實施了嚴格的保護措施。經過多年的努力,加州紅腿蛙的種群在一些地區已經開始恢復,雖然仍面臨挑戰,但這個案例展示了綜合性保育策略的有效性。
在亞洲,諸羅樹蛙(Rhacophorus arvalis)的保育故事也值得關注。這種樹蛙是台灣的特有種,由於棲息地破壞和污染,其數量曾經急劇下降。為了保護這個物種,台灣的科學家和保育人士展開了一系列行動。他們不僅進行了詳細的生態學研究,還開展了棲息地恢復工作。特別是,他們在諸羅樹蛙的棲息地周圍建立了保護區,並與當地社區合作,推廣環境友好型的農業實踐。這些努力使得諸羅樹蛙的種群在一些地區得到了恢復,成為台灣本土物種保育的一個成功範例。
這些成功案例展示了多種保育策略的有效性。人工繁育是許多案例中的關鍵環節,它為瀕危物種提供了一個安全網,確保了基因多樣性的保存。同時,棲息地保護和恢復也是不可或缺的,因為只有在適宜的自然環境中,這些物種才能長期生存。此外,疾病管理、入侵物種控制等針對性措施也在許多案例中發揮了重要作用。
值得注意的是,這些成功案例大多涉及長期的努力和多方面的合作。政府部門、科研機構、保育組織和當地社區的共同參與是成功的關鍵。同時,公眾教育和參與也在許多案例中發揮了重要作用,提高了人們對蛙類保護的認識和支持。
這些成功案例不僅挽救了特定的蛙類物種,還對整個生態系統產生了積極影響。蛙類作為生態系統中的重要一環,其恢復有助於維持生態平衡,同時也為其他依賴於相同棲息地的物種帶來了益處。
然而,我們也必須認識到,這些成功案例並不意味著保育工作的結束。許多恢復的種群仍然脆弱,需要持續的保護和監測。同時,這些經驗也需要被推廣和應用到其他瀕危蛙類的保育工作中,以應對全球範圍內蛙類面臨的持續威脅。
13. 蛙類與人類
蛙類與人類的關係由來已久,深入到文化、經濟、科學和生態等多個層面。在許多文化中,蛙類常被視為吉祥或神聖的象徵,如古埃及將蛙神赫克特視為生育和重生的象徵。在中國傳統文化中,蛙被視為招財進寶的吉祥物。
在飲食文化方面,蛙類在全球多個地區被視為美食。法國的蛙腿就是著名的美食之一,而在東南亞地區,如越南和泰國,炒田雞也是常見的菜餚。這種飲食習慣促進了蛙類養殖業的發展,成為一些地區的重要經濟來源。
在科學研究領域,蛙類扮演著重要角色。它們常被用作生物學研究的模型生物,特別是在發育生物學和生理學研究中。例如,非洲爪蟾(Xenopus laevis)因其卵細胞大且易於操作,成為研究細胞分裂和早期胚胎發育的理想對象。
在醫學方面,蛙類皮膚分泌物中的一些化合物被發現具有抗菌、麻醉等藥用價值,正成為新藥研發的重要來源。同時,蛙類作為環境健康的指標生物,在生態監測和環境保護中發揮著重要作用。
近年來,隨著環境意識的提高,蛙類保育已成為全球關注的話題。許多地方開展了蛙類觀察活動,既提高了公眾的環保意識,也促進了生態旅遊的發展。然而,過度捕捉和棲息地破壞仍然威脅著許多蛙類物種,需要我們在利用與保護之間尋找平衡。
13.1 蛙類作為烹飪美食
蛙類作為烹飪美食在全球多個地區都有悠久的傳統,其獨特的口感和營養價值使其成為許多美食愛好者追捧的對象。在不同的文化中,蛙類料理展現出豐富多樣的烹飪方式和風味特色。
在西方國家中,法國可能是最著名的蛙類美食愛好者。法式蛙腿(Cuisses de grenouille)是一道備受推崇的傳統菜餚,通常採用牛蛙或水蛙的後腿部分。烹製方法多樣,最常見的是將蛙腿裹上麵粉後油炸,再搭配蒜蓉、歐芹和檸檬汁調味。這道菜以其嫩滑的口感和淡淡的雞肉味而聞名,被認為是法國南部普羅旺斯地區的特色美食。
在亞洲,蛙類料理更是種類繁多。中國的粵菜中就有多道著名的蛙類菜餚。例如,高檔粵菜餐廳常見的「飛天蛙」,是將牛蛙去骨後填入魚漿,再裹上麵衣炸至金黃,口感鮮嫩多汁。另一道名菜「雪蛙鮮奶」則是將蛙肉與鮮奶、蛋白一起烹製,口感滑嫩,味道清淡鮮美。在四川菜系中,「水煮牛蛙」以其麻辣鮮香的口味深受歡迎,牛蛙肉質鮮嫩,搭配麻辣湯底,風味獨特。
泰國的蛙類料理同樣豐富多樣。泰式炒蛙(Pad Kob)是一道常見的街頭小吃,將蛙肉與辣椒、蒜、羅勒等香料一起快炒,味道辛辣鮮香。另一道名為「蛙肉冬陰功」的菜餚,將蛙肉加入到泰國著名的酸辣湯中,湯汁酸辣開胃,蛙肉吸收湯汁的味道後更顯鮮美。
在越南,蛙類也是受歡迎的食材。越式炸蛙腿(Đùi ếch chiên)是一道常見的下酒菜,蛙腿裹上特製的調味麵糊後油炸,外酥內嫩,常搭配魚露調味的醬汁食用。另一道名為「蓮葉烤蛙」的菜餚,將調味後的整隻蛙包裹在蓮葉中烤製,蛙肉吸收了蓮葉的清香,口感獨特。
在印度尼西亞,特別是在爪哇島,蛙類料理也頗受歡迎。一道名為「Swike」的菜餚,是將蛙肉與多種香料如薑黃、南薑、蒜等一起燉煮,湯汁濃郁,蛙肉味道豐富。
回到歐洲,除了法國,西班牙的某些地區也有食用蛙類的傳統。在西班牙的拉里奧哈地區,有一道名為「Ancas de rana al pil-pil」的菜餚,將蛙腿與大蒜、辣椒一起在橄欖油中慢煎,味道鮮美,口感酥脆。
值得注意的是,隨著人們對蛙類料理的需求增加,蛙類養殖業也隨之發展。中國是世界上最大的食用蛙養殖國,主要養殖品種包括牛蛙、虎紋蛙等。這些養殖蛙不僅供應國內市場,還大量出口到其他國家。
從營養學角度來看,蛙肉被認為是一種優質的蛋白質來源,脂肪含量較低,並富含維生素和礦物質。特別是蛙腿肉,其蛋白質含量高,脂肪含量低,被視為一種健康的肉類選擇。
然而,我們也必須認識到,過度捕捉野生蛙類可能對生態系統造成嚴重影響。許多國家已經開始限制野生蛙類的捕捉,轉而鼓勵養殖。同時,一些環保組織也呼籲消費者在選擇蛙類食品時,優先考慮可持續來源的產品。
在烹飪過程中,新鮮度是決定蛙類料理品質的關鍵因素。由於蛙肉容易變質,通常建議購買活蛙或冰鮮蛙肉。在處理時,需要特別注意衛生,以避免可能的寄生蟲感染。
蛙類料理的烹飪技巧也值得關注。由於蛙肉質地較為嬌嫩,容易過度烹煮變得乾柴,因此控制火候和烹飪時間至關重要。許多廚師建議採用快速烹調的方法,如煎炒或油炸,以保持肉質的鮮嫩多汁。
隨著全球化的發展,蛙類料理也在不斷創新。一些現代廚師嘗試將傳統的蛙類烹飪方法與新的烹飪技術和食材結合,創造出新的口味和呈現方式。例如,有些廚師嘗試將蛙肉製成刺身或塔塔,以全新的方式展現蛙肉的鮮美。
儘管蛙類美食在許多地方備受推崇,但在一些文化中,食用蛙類仍然是一個有爭議的話題。一些人基於宗教、文化或個人信仰的原因,可能會避免食用蛙類。因此,在不同文化背景下提供和享用蛙類料理時,需要尊重和理解這些差異。
13.2人類蛙類養殖與經濟效益
人類蛙類養殖是一個迅速發展的產業,不僅為滿足食用需求提供了穩定的來源,還創造了可觀的經濟效益。這一產業在全球多個地區都有開展,但以亞洲和美洲最為普遍。蛙類養殖不僅為農民提供了新的收入來源,還在一定程度上減輕了對野生蛙類資源的壓力。
在蛙類養殖中,最常見的品種是牛蛙(Rana catesbeiana),這種原產於北美的蛙類因其體型大、生長快、適應性強而成為養殖的首選。除了牛蛙,虎紋蛙(Hoplobatrachus tigerinus)和青蛙(Rana nigromaculata)等品種也在不同地區得到廣泛養殖。
中國是世界上最大的蛙類養殖國,年產量占全球總產量的百分之八十以上。中國的蛙類養殖主要集中在廣東、四川、湖南等省份。這些地區氣候適宜,水資源豐富,為蛙類養殖提供了理想的自然條件。中國的蛙類養殖模式多樣,包括池塘養殖、稻田養殖和工廠化養殖等。其中,稻田養殖模式被認為是一種生態友好的養殖方式,蛙類可以在稻田中捕食害蟲,減少農藥使用,同時蛙糞可以作為天然肥料滋養稻田,形成了一個良性的生態循環。
在東南亞地區,如泰國、越南和印尼,蛙類養殖也是一個重要的經濟活動。這些國家的氣候條件適合蛙類生長,同時當地有食用蛙類的傳統,為養殖業提供了穩定的市場需求。泰國的蛙類養殖業特別發達,除了滿足國內市場,還大量出口到歐美國家。
在美洲,巴西是蛙類養殖的主要國家之一。巴西的蛙類養殖始於20世紀70年代,主要養殖品種是牛蛙。巴西的養殖技術較為先進,許多養殖場採用了溫室養殖系統,可以全年生產。巴西的蛙肉產品不僅供應國內市場,還出口到美國、法國等國家。
蛙類養殖的經濟效益主要體現在以下幾個方面:首先,蛙肉作為一種高蛋白、低脂肪的食材,在市場上有較高的售價。以中國市場為例,養殖蛙的價格通常是普通養殖魚類的2-3倍。其次,蛙類的生長周期較短,通常在3-6個月就可以達到上市規格,這意味著資金周轉速度快,經濟效益顯著。
此外,蛙類養殖還衍生出了多個相關產業,進一步擴大了其經濟效益。例如,蛙皮可以用於製作皮革製品,蛙油被用於化妝品生產,蛙的內臟可以製作成動物飼料。在中國,甚至還開發出了蛙類保健品市場,聲稱蛙類提取物具有增強免疫力等功效。
然而,蛙類養殖也面臨著一些挑戰。首先是疾病防控問題。密集養殖環境容易導致疾病的快速傳播,特別是壺菌病(Chytridiomycosis)等真菌感染,可能會造成大規模的死亡。為此,養殖戶需要投入大量資金用於疾病防控和水質管理。
其次是環境問題。一些不規範的養殖場可能會造成水體污染,影響周邊環境。此外,如果養殖的外來種如牛蛙逃逸到野外,可能會對當地生態系統造成破壞。因此,許多國家開始實施嚴格的養殖管理制度,要求養殖戶採取措施防止蛙類逃逸。
蛙類養殖的技術也在不斷進步。例如,中國的一些養殖場開始採用循環水養殖系統,不僅節約了水資源,還提高了養殖密度和產量。同時,通過改良飼料配方,養殖戶可以加快蛙類的生長速度,提高肉質品質。
在經濟效益方面,蛙類養殖為農村地區創造了大量就業機會。以中國為例,蛙類養殖產業已經形成了一個完整的產業鏈,包括苗種繁育、飼料生產、養殖、加工和銷售等環節,為數十萬人提供了就業崗位。
此外,蛙類養殖還帶動了相關產業的發展。例如,為了滿足養殖需求,專門的蛙類飼料生產企業應運而生。同時,蛙類加工業也在快速發展,生產各種冷凍蛙肉產品和即食蛙類食品,增加了產品附加值。
值得注意的是,隨著人們對食品安全和環境保護意識的提高,有機蛙類養殖開始受到關注。這種養殖方式不使用抗生素和化學添加劑,雖然成本較高,但產品可以賣出更高的價格,迎合了高端市場的需求。
儘管蛙類養殖帶來了顯著的經濟效益,但我們也需要關注其可能帶來的生態風險。一些國家已經開始實施嚴格的養殖許可制度,要求養殖戶採取措施防止養殖蛙類逃逸到野外。同時,鼓勵養殖本土蛙種,減少對生態系統的潛在威脅。
13.3 蛙類在科學研究中的應用
蛙類在科學研究中扮演著極其重要的角色,其獨特的生理特征和生活史特點使其成為多個科學領域的理想研究對象。從基礎生物學到醫學應用,蛙類為科學家們提供了豐富的研究素材和寶貴的見解。
在發育生物學領域,非洲爪蟾(Xenopus laevis)是最廣泛使用的模式生物之一。這種蛙類的卵細胞特別大,易於觀察和操作,使其成為研究早期胚胎發育過程的理想對象。科學家們利用非洲爪蟾研究了細胞分裂、組織分化和器官形成等關鍵發育過程。例如,著名的脊椎動物發育生物學家約翰·格登(John Gurdon)利用非洲爪蟾進行了核移植實驗,為克隆技術的發展奠定了基礎,並因此獲得了2012年諾貝爾生理學或醫學獎。
在生理學研究中,蛙類也發揮了重要作用。蛙類的皮膚具有獨特的滲透性和呼吸功能,成為研究離子通道和膜運輸的理想模型。科學家們利用蛙皮研究了水分和電解質平衡的調節機制,這些研究不僅增進了我們對蛙類生理的理解,也為人類醫學提供了重要參考。此外,蛙類的心臟結構相對簡單,易於觀察和實驗,因此常被用於心血管生理學研究。
在神經生物學領域,蛙類的神經系統相對簡單但功能完整,為研究神經信號傳導和行為控制提供了理想的模型。例如,科學家們利用蛙類研究了神經肌肉接頭的結構和功能,這些研究對理解人類神經系統疾病如肌無力症等具有重要意義。
蛙類在毒理學研究中也扮演著重要角色。由於蛙類對環境變化特別敏感,它們常被用作環境污染的生物指標。科學家們通過觀察蛙類的生長、發育和行為變化,可以評估水體和土壤中污染物的影響。例如,一些研究發現某些農藥和工業污染物會導致蛙類畸形率增加,這些發現引起了人們對環境保護的重視。
在進化生物學領域,蛙類的多樣性為研究適應性輻射和物種形成提供了豐富的素材。科學家們通過比較不同蛙類物種的形態、生理和行為特征,探討了生物如何適應不同的環境。例如,對樹蛙的研究揭示了它們如何進化出適應樹棲生活的特殊結構,如吸盤狀的腳趾。
蛙類在再生生物學研究中也有重要應用。某些蛙類物種具有強大的再生能力,能夠重新生長失去的四肢或尾巴。科學家們通過研究這些再生過程,希望能為人類再生醫學提供新的思路。例如,對墨西哥蠑螈(雖然不是蛙類,但也是兩棲類)的研究為理解組織再生的分子機制提供了重要線索。
在藥物研究領域,蛙類皮膚分泌物中的生物活性物質引起了科學家們的廣泛興趣。許多蛙類物種能夠分泌具有抗菌、抗病毒或鎮痛作用的化合物,這些物質為新藥開發提供了潛在的來源。例如,從南美樹蛙皮膚中提取的一種肽被發現具有強大的鎮痛作用,其效力甚至超過嗎啡,但不具有成癮性,這一發現為開發新型止痛藥物開闢了新的方向。
在行為生態學研究中,蛙類的繁殖行為和聲音通訊為科學家們提供了豐富的研究素材。通過研究蛙類的求偶行為和鳴叫模式,科學家們探討了動物通訊和性選擇的演化機制。例如,對綠樹蛙的研究揭示了雌性如何通過雄性的鳴叫來選擇配偶,這些發現豐富了我們對動物行為進化的理解。
蛙類在基因組學研究中也發揮了重要作用。隨著測序技術的進步,越來越多的蛙類基因組被解析。這些基因組數據不僅有助於我們理解蛙類的進化歷史,也為比較基因組學研究提供了寶貴的資源。例如,通過比較不同蛙類物種的基因組,科學家們發現了與適應性進化相關的基因變異。
在生物醫學研究中,蛙類也有重要應用。例如,某些蛙類的皮膚分泌物被發現具有抗癌潛力。科學家們正在研究這些化合物的作用機制,希望能為癌症治療提供新的思路。此外,蛙類的免疫系統研究也為人類免疫學研究提供了重要參考。
蛙類在環境科學研究中的應用尤為重要。作為生態系統中的關鍵物種,蛙類的健康狀況可以反映整個生態系統的健康。科學家們通過監測蛙類種群的變化,評估氣候變化、棲息地破壞等因素對生態系統的影響。這些研究不僅有助於保護蛙類本身,也為制定更廣泛的環境保護策略提供了科學依據。
儘管蛙類在科學研究中的應用如此廣泛,但我們也需要注意到使用蛙類進行研究時的倫理問題。科學界正在努力發展替代方法,如體外細胞培養和計算機模擬,以減少對活體蛙類的使用。同時,對於必須使用活體蛙類的研究,科學家們也在不斷完善實驗規範,以確保動物福利。
13.4 蛙類在傳統醫學中的應用
蛙類在傳統醫學中的應用由來已久,這種應用涵蓋了多個文化和地區,反映了人類對自然資源的利用以及對健康的追求。在中國傳統醫學中,蛙類被視為具有多種藥用價值的動物。其中最為人所知的可能是蟾蜍,它的皮膚分泌物被稱為「蟾酥」,被認為具有解毒、消腫、鎮痛的功效。中醫典籍中記載,蟾酥可用於治療癰腫、喉痹、牙痛等症狀。另一種常見的藥用蛙類是雪蛤,它實際上是東北林蛙的卵巢,被認為具有滋陰補腎、潤肺止咳的功效,常被製成膏狀食用。
在東南亞地區,如越南和柬埔寨,某些蛙類被用於製作傳統藥酒。人們相信這種藥酒能夠增強體力、改善關節問題,甚至提高性功能。印度的阿育吠陀醫學中也有使用蛙類的記載,某些蛙類被用於治療哮喘、水腫等疾病。在南美洲,尤其是亞馬遜地區,原住民長期使用某些樹蛙的皮膚分泌物作為狩獵時的箭毒,這些毒素後來被發現具有潛在的醫療價用,如痛覺抑制劑。
然而,需要注意的是,傳統醫學中使用蛙類的做法往往缺乏現代科學的嚴謹驗證。許多聲稱的療效可能源於文化傳統或個人經驗,而非系統性的臨床研究。更重要的是,某些做法可能對蛙類種群造成嚴重威脅。例如,為了獲取藥用成分而過度捕捉野生蛙類,可能導致局部種群數量驟減,甚至滅絕。這種情況在某些珍稀蛙類身上尤為明顯,如金蛙(蝦蟆油)的濫捕就曾導致其瀕臨絕種。
現代醫學研究已經開始關注某些蛙類分泌物中的生物活性物質。例如,從某些樹蛙皮膚中提取的肽類化合物被發現具有抗菌、抗病毒甚至抗癌的潛力。這些研究為傳統醫學中蛙類應用提供了一定的科學基礎,同時也開闢了新的藥物研發方向。然而,這類研究通常採用生物技術手段,如基因工程或體外培養,而不是直接使用野生蛙類,這種方法既能保護野生種群,又能確保產品的安全性和穩定性。
在台灣,雖然傳統上也有使用蛙類入藥的做法,但隨著現代醫學的發展和保育意識的提高,這種做法已經大幅減少。目前,台灣更多地關注蛙類的生態價值和保育需求,而非其潛在的藥用價值。這種轉變反映了現代社會對生物多樣性保護的重視,以及對傳統做法的反思。
儘管如此,蛙類在傳統醫學中的應用仍然是一個值得深入研究的領域。一方面,我們需要用現代科學方法來驗證傳統知識,篩選出真正有效且安全的應用;另一方面,我們也需要平衡醫藥需求和生態保護,確保任何利用都建立在可持續的基礎上。這需要醫學、生態學、保育生物學等多個領域的專家共同努力,制定合理的利用策略和保護措施。
此外,蛙類在傳統醫學中的應用也引發了一系列倫理問題。我們需要考慮,為了人類健康而大量捕捉或飼養蛙類是否合乎道德?如何在尊重傳統文化和保護瀕危物種之間找到平衡?這些問題沒有簡單的答案,需要社會各界深入討論和權衡。
13.5 蛙類作為人類寵物
蛙類作為寵物在許多國家和地區逐漸成為一種流行趨勢,這種現象反映了人們對自然的嚮往和對獨特寵物的追求。相比傳統的貓狗寵物,蛙類具有獨特的魅力和挑戰。它們的外觀多樣,從鮮豔奪目到擬態隱蔽,能為飼養者帶來視覺上的享受。同時,蛙類的生活習性和行為模式也頗具觀賞性,例如它們捕食昆蟲的敏捷動作,或是在繁殖季節的鳴叫,都能為飼養者提供獨特的體驗。
然而,將蛙類作為寵物飼養也面臨諸多挑戰和爭議。首先是環境需求的複雜性。不同種類的蛙類對溫度、濕度和棲息地的要求各不相同,有些需要模擬熱帶雨林環境,有些則需要乾燥的沙漠地形。這就要求飼養者具備相當的知識和技能,能夠為蛙類創造適宜的生存環境。例如,樹蛙類需要較高的濕度和攀爬的枝條,而陸生蛙類則需要較乾燥的環境和隱蔽處。
飲食是另一個需要特別關注的問題。大多數蛙類是肉食性動物,主要以昆蟲為食。在家庭環境中,飼養者需要定期提供活的食物,如蟋蟀、蠐螬或果蠅等。這不僅增加了飼養的複雜度,也可能對一些飼養者造成心理負擔。此外,確保食物的營養均衡也是一個挑戰,飼養者需要了解不同蛙類的營養需求,並可能需要添加補充劑以確保寵物的健康。
蛙類的健康管理也是一個重要課題。它們對環境變化非常敏感,容易受到應激和疾病的影響。常見的健康問題包括真菌感染、寄生蟲和營養不良等。飼養者需要定期觀察蛙類的行為和外觀,及時發現問題並尋求專業獸醫的幫助。然而,相比貓狗等常見寵物,專門治療兩棲動物的獸醫相對較少,這可能增加了飼養的難度。
從生態保護的角度來看,將蛙類作為寵物飼養也引發了一些爭議。野生蛙類的捕捉可能對自然種群造成壓力,特別是對於一些稀有或瀕危物種。雖然現在市面上的寵物蛙大多來自人工繁殖,但仍有非法捕捉和交易的情況發生。此外,飼養者如果不小心或故意將外來蛙類釋放到野外,可能會引入入侵物種,對本地生態系統造成嚴重影響。例如,在台灣,外來的斑腿樹蛙就因為寵物逃逸或故意釋放而成為入侵物種,對本地蛙類構成威脅。
法律和倫理問題也是不容忽視的方面。不同國家和地區對於蛙類寵物的管理政策各不相同。有些地方嚴格限制某些蛙類物種的飼養和交易,而有些地方則相對寬鬆。飼養者需要了解並遵守當地的相關法規。同時,飼養蛙類作為寵物也引發了一些倫理討論,例如是否應該將野生動物馴化為寵物,以及如何在滿足人類需求和尊重動物權利之間取得平衡。
儘管存在這些挑戰和爭議,但對於那些願意投入時間和精力的愛好者來說,飼養蛙類作為寵物仍然可以是一種有趣且有教育意義的體驗。許多飼養者通過照顧這些兩棲動物,加深了對自然界的理解和欣賞。一些學校和教育機構也會飼養蛙類作為教學工具,幫助學生了解生物多樣性和生態系統。
為了負責任地飼養蛙類寵物,有幾點建議值得注意。首先,飼養者應該選擇適合自己能力和條件的蛙類物種。對於初學者來說,諸如美洲綠樹蛙或非洲爪蟾等較為容易飼養的物種可能是不錯的選擇。其次,盡可能從可靠的來源購買人工繁殖的個體,避免參與野生動物的非法交易。再者,飼養者應該充分了解所選蛙類的生活習性和需求,並準備好適當的飼養環境。這包括合適大小的容器、適當的底材、溫度和濕度控制設備等。
此外,飼養者還需要注意個人衛生。許多兩棲動物可能攜帶沙門氏菌等病原體,因此在接觸蛙類或清理其生活環境後,必須徹底洗手。同時,避免將蛙類放在食品準備區域或兒童可能接觸到的地方也很重要。
對於那些決定不再飼養蛙類寵物的人,正確的處理方式至關重要。無論如何都不應該將它們釋放到野外,這不僅可能危及蛙類本身的生存,也可能對本地生態系統造成破壞。相反,應該尋求專業機構的幫助,如動物收容所或專門的爬蟲類和兩棲類救援組織。
總的來說,將蛙類作為寵物飼養是一項需要深思熟慮的決定。它需要飼養者具備足夠的知識、耐心和責任感。當這些條件得到滿足時,飼養蛙類不僅可以為人們帶來樂趣,還能增進對自然世界的理解和尊重。然而,我們也必須時刻牢記,即使是作為寵物,蛙類仍然是野生動物,它們的福祉和對生態系統的潛在影響都應該得到認真考慮。
13.6 受蛙類啟發的科技
蛙類作為演化歷程中的成功物種,其獨特的生理結構和行為特徵為科學家和工程師提供了豐富的靈感來源,促進了多個領域的技術創新。這些受蛙類啟發的科技涵蓋了從材料科學到機器人技術等廣泛範疇,展現了生物仿生學在現代科技發展中的重要地位。
在材料科學領域,蛙類的皮膚結構引發了諸多創新。許多蛙類具有特殊的皮膚特性,能夠在乾燥和潮濕環境中自如轉換。科研人員受此啟發,開發出了具有類似特性的智能材料。例如,某些研究團隊設計出了模仿蛙皮的自潤滑表面,這種表面能夠在乾燥時保持光滑,遇水時則會形成一層保護膜。這種技術可能應用於自清潔玻璃、防霧鏡片等產品,大大提高了材料的實用性和耐用性。
蛙類的黏附能力也是科技創新的重要靈感來源。樹蛙能夠在光滑垂直表面上自如攀爬,這種能力源於其腳掌上的特殊結構。科學家通過研究樹蛙腳掌的微觀結構,開發出了新型的黏附材料。這些材料不僅具有強大的黏附力,還可以重複使用且不留殘留物。這項技術可能徹底改變多個領域,從醫療器械到工業應用,甚至可能用於太空探索中的無重力環境操作。
在機器人領域,蛙類的跳躍能力為研究人員提供了寶貴的參考。蛙類能夠利用後腿肌肉儲存能量,然後瞬間釋放,實現遠距離跳躍。受此啟發,工程師們設計出了能夠模仿蛙類跳躍的機器人。這些機器人不僅能夠實現高效的運動,還可以適應複雜的地形。這種技術在搜救、探測等領域有著廣闊的應用前景,特別是在人類難以到達的危險或狹小環境中。
蛙類的感官系統也為科技創新提供了重要啟示。例如,某些蛙類具有極其敏銳的聽覺,能夠在嘈雜的環境中精確定位聲音來源。研究人員通過研究蛙類的聽覺系統,開發出了新型的聲音定位技術。這種技術可能應用於助聽器的改進,提高在複雜聲音環境中的語音辨識能力。此外,這種技術還可能用於水下聲納系統的優化,提高海洋探測和通信的效率。
在光學領域,某些蛙類的眼睛結構也引發了科學家的興趣。例如,有些樹蛙具有能夠在黑暗中高效捕捉光線的特殊視網膜結構。研究人員通過模仿這種結構,開發出了新型的光敏感器。這種感器在弱光環境下表現優異,可能用於改進夜視設備或提高數碼相機在低光條件下的成像質量。
蛙類的變色能力也為科技創新提供了思路。某些蛙類能夠根據環境改變體色,這種能力源於其皮膚中的特殊色素細胞。科學家受此啟發,開發出了能夠根據環境變化顏色的智能材料。這種材料可能用於製作迷彩服,提高軍事裝備的隱蔽性。在民用領域,這種技術可能應用於智能窗戶,根據光線強度自動調節透光度,提高建築物的能源效率。
在醫療科技領域,蛙類的某些生理特性也為研究人員提供了寶貴的靈感。例如,某些蛙類能夠在極端寒冷的環境中存活,其體內會產生一種特殊的抗凍蛋白。科學家通過研究這種蛋白質的結構和功能,開發出了新型的器官保存液。這種技術可能大大延長器官移植過程中的器官保存時間,提高移植手術的成功率。
蛙類的再生能力也是醫學研究的重要參考。某些蛙類幼體具有再生斷肢的能力,這一特性引發了再生醫學領域的大量研究。科學家們希望通過深入了解蛙類的再生機制,為人類器官再生和傷口癒合提供新的治療方法。雖然這一領域的研究仍處於早期階段,但已經在組織工程和幹細胞治療等方面取得了一些進展。
在環境監測領域,蛙類的生理特性也被用作發展新技術的靈感來源。眾所周知,蛙類對環境變化極為敏感,常被視為生態系統健康的指標物種。研究人員基於這一特性,開發出了模仿蛙類皮膚的環境感應器。這種感應器能夠檢測水質、空氣質量等環境參數的微小變化,為環境監測提供了新的工具。
蛙類的鳴叫行為也啟發了聲學技術的發展。某些蛙類能夠在嘈雜的環境中精確傳遞聲音信息,這種能力引起了通信工程師的興趣。通過研究蛙類的發聲機制和聲音傳播模式,研究人員開發出了新型的聲學編碼技術,可能用於提高在複雜環境下的通信質量。
值得注意的是,這些受蛙類啟發的科技創新不僅展示了自然界的奇妙,也凸顯了保護生物多樣性的重要性。每一種生物都可能蘊含著未被發現的科技潛力,而蛙類及其棲息地的保護不僅對生態平衡至關重要,也可能為未來的科技突破提供關鍵靈感。
這些創新技術的發展過程也強調了跨學科合作的重要性。從生物學家到工程師,從材料科學家到醫學研究人員,不同領域的專家通力合作,將自然界的奇妙轉化為實用的技術解決方案。這種跨學科的研究方法不僅推動了科技創新,也深化了我們對自然世界的理解。
13.7 蛙類研究與保育的倫理考量
蛙類研究與保育涉及複雜的倫理考量,這些考量反映了科學進步、生態保護和人類需求之間的微妙平衡。隨著人類對蛙類及其棲息地影響的加深,以及科學研究方法的不斷演進,相關的倫理問題變得越發突出和多元化。
在科學研究領域,使用蛙類作為實驗對象一直是生物學和醫學研究的重要部分。然而,這種做法也引發了動物福利的倫理爭議。研究人員必須權衡科學發現的潛在價值與實驗動物所承受的痛苦。例如,在研究蛙類的再生能力時,可能需要對實驗對象進行截肢或組織損傷。儘管這些研究可能帶來重要的醫學突破,但也不可避免地給實驗動物造成痛苦。因此,許多研究機構已經建立了嚴格的動物實驗倫理審查制度,要求研究者詳細說明實驗的必要性,並採取措施最大限度地減少動物痛苦。
此外,野外研究中的取樣和標記技術也引發了倫理討論。為了追蹤蛙類的行為和遷移模式,研究人員可能需要對野生個體進行標記,如使用腳趾切除法或植入微型晶片。這些方法雖然能提供寶貴的數據,但可能對個體造成永久性傷害或影響其生存能力。因此,研究者必須仔細評估研究的價值與對個體和種群的潛在影響,並尋求更加人道的替代方法,如使用特殊顏料進行臨時標記。
在保育實踐中,人工繁殖和再引入項目也面臨著複雜的倫理問題。這些項目旨在保護瀕危蛙類,但可能帶來意想不到的生態後果。例如,人工繁殖可能導致遺傳多樣性的減少,影響種群的長期生存能力。再引入項目則可能干擾現有的生態平衡,或者將疾病引入野生種群。因此,保育人員必須謹慎評估這些項目的長期影響,並制定嚴格的監測和管理計劃。
蛙類保育還涉及資源分配的倫理問題。在有限的保育資金和人力資源下,如何決定優先保護哪些物種或棲息地?是否應該將資源集中於明星物種,還是平均分配給所有瀕危物種?這些決策不僅涉及科學考量,還涉及道德和價值判斷。例如,某些蛙類可能在生態系統中扮演關鍵角色,其保護可能帶來更廣泛的生態效益。但同時,我們也有道德義務保護所有瀕危物種,即使它們可能不那麼「重要」或「引人注目」。
在應對氣候變化對蛙類的影響時,也存在倫理困境。隨著全球氣溫上升和極端天氣事件增加,許多蛙類面臨棲息地喪失的威脅。一些保育者提出了「輔助遷徙」的概念,即人為將物種遷移到更適合的新棲息地。然而,這種做法可能干擾接收地的生態系統,引發新的倫理爭議。我們是否有權利為了拯救一個物種而潛在地危及另一個生態系統?這個問題沒有簡單的答案,需要權衡多方面的考慮。
蛙類研究中的基因編輯技術應用也帶來了新的倫理挑戰。例如,科學家們正在探索使用基因編輯技術來增強蛙類對某些疾病的抵抗力。雖然這可能有助於保護瀕危物種,但也引發了對基因改造生物釋放到自然環境中的擔憂。我們需要仔細評估這種技術的長期生態影響和潛在風險。
在處理入侵蛙類物種時,研究者和保育者面臨著另一個倫理困境。為了保護本地生態系統,有時需要大規模清除入侵物種。然而,這種做法可能被視為殘忍或不人道,特別是當涉及到大量個體的處置時。保育者需要在生態系統健康和動物福利之間尋求平衡,同時也要考慮公眾對這些行動的看法和接受度。
蛙類研究和保育還涉及文化敏感性的問題。在某些文化中,蛙類可能具有特殊的文化或宗教意義。研究者和保育者在開展工作時,需要尊重當地的文化傳統和信仰。例如,在某些地區,特定的蛙類可能被視為神聖或具有藥用價值。在這種情況下,科學研究和保育工作需要與當地社區密切合作,尋求共識和互利的解決方案。
此外,蛙類研究和保育項目的資金來源也可能引發倫理爭議。當資金來自可能對環境造成負面影響的企業或行業時,研究者和保育者可能面臨利益衝突。他們需要權衡接受這些資金的潛在好處與可能的負面影響,並確保研究的獨立性和客觀性不受損害。
知識共享和數據管理也是蛙類研究和保育中的重要倫理考量。研究者有責任公開分享他們的發現,以促進科學進步和保育努力。然而,這也需要平衡保護敏感信息的需求,例如瀕危物種的精確位置,以防止非法捕獵或干擾。
最後,蛙類研究和保育的倫理考量還延伸到公眾教育和參與的領域。研究者和保育者有責任向公眾傳播準確的信息,提高人們對蛙類重要性的認識。然而,他們也需要謹慎處理可能引發恐慌或誤解的信息,如某些蛙類作為疾病載體的角色。
綜上所述,蛙類研究與保育的倫理考量涉及多個層面,從個體動物福利到生態系統健康,從文化敏感性到資源分配。這些考量反映了科學、保育和社會價值觀之間的複雜互動。研究者和保育者需要不斷反思和評估他們的做法,確保在追求科學知識和保護生物多樣性的同時,也尊重倫理原則和社會責任。這需要跨學科的合作,包括生物學家、生態學家、倫理學家和社會科學家的共同努力,以制定全面的倫理指南和最佳實踐。同時,這也要求我們在更廣泛的社會背景下思考蛙類研究和保育的角色,平衡科學進步、生態保護和人類需求。
14. 蛙類保育與研究
蛙類研究與保育的未來方向是一個廣闊而複雜的領域,涉及多個方面的創新和挑戰。隨著全球環境變化和人類活動的加劇,蛙類面臨著前所未有的威脅,這促使科學家和保育者不斷探索新的研究方法和保護策略。
在蛙類新興保育策略方面,我們看到了一些令人振奮的發展。其中之一是整合性棲息地管理方法的採用。這種方法不僅關注單一物種的保護,而是致力於維護整個生態系統的健康。例如,在澳大利亞,研究人員正在實施一項綜合性的濕地恢復計劃,旨在同時保護多個蛙類物種及其共生物種。這個項目不僅包括棲息地的物理恢復,還考慮了水質改善、外來物種控制以及周邊土地使用的管理。這種全面的方法能夠為蛙類創造更加穩定和多樣化的生存環境,提高其長期生存的機會。
另一個新興的保育策略是疾病管理。近年來,壺菌病(Chytridiomycosis)已成為全球蛙類面臨的主要威脅之一。為應對這一挑戰,科學家們正在開發創新的方法。例如,一些研究團隊正在探索使用益生菌來增強蛙類對壺菌的抵抗力。在巴拿馬,研究人員已經成功地將帶有有益細菌的蛙類重新引入到野外,這些蛙類表現出更強的疾病抵抗能力。這種生物防禦方法不僅可能有效對抗壺菌病,還可能應用於其他蛙類疾病的預防和控制。
此外,氣候變化適應策略也成為蛙類保育的重要方向。隨著全球氣溫上升,許多蛙類面臨著棲息地丟失或不適宜的風險。為此,科學家們正在研究如何幫助蛙類適應氣候變化。一種方法是創建氣候避難所,即在預計未來氣候條件下仍然適合蛙類生存的區域。例如,在美國加州,研究人員正在識別和保護高海拔地區的某些水體,這些地方可能成為當地蛙類在氣候變暖時的重要避難所。另一種策略是協助遷移,即將蛙類遷移到更適合的新棲息地。雖然這種方法存在爭議,但在某些情況下可能是拯救極度瀕危物種的唯一選擇。
在兩棲類的恢復生態學方面,我們看到了一些創新的方法和實踐。恢復生態學不僅關注物種的直接保護,還致力於重建和修復已經退化的生態系統。在蛙類保育中,這通常涉及濕地和水生環境的恢復。
一個典型的例子是在歐洲進行的泥炭地恢復項目。泥炭地是許多蛙類的重要棲息地,但在過去幾個世紀中遭受了嚴重破壞。在英國和愛爾蘭,大規模的泥炭地恢復工程正在進行,這不僅有助於蛙類保護,還能提供碳封存等生態系統服務。這些項目包括重新灌水、植被重建以及控制侵入性植物等措施。初步結果顯示,一些本地蛙類種群在恢復的泥炭地中開始恢復。
另一個重要的恢復生態學應用是城市生態系統的改造。隨著城市化的加速,許多蛙類失去了原有的棲息地。為此,一些城市正在實施創新的綠色基礎設施計劃,如建造雨水花園、生態廊道和人工濕地。這些設施不僅能為城市蛙類提供棲息地,還能改善水質和減少城市熱島效應。例如,在新加坡,政府正在推行"公園連接器"項目,通過綠色走廊將城市公園連接起來,為包括蛙類在內的野生動物提供移動通道。
在恢復生態學中,一個新興的趨勢是採用"重建生態系統"的方法。這種方法不僅僅是恢復原有的生態系統,而是創造能夠在當前和未來環境條件下蓬勃發展的新生態系統。這對於蛙類保育特別重要,因為許多原有的棲息地可能已經無法在氣候變化的背景下維持。例如,在澳大利亞的某些地區,科學家們正在設計和建造能夠適應更乾燥氣候的人工濕地,這些濕地包含了深水區和季節性淹水區,可以為不同的蛙類物種提供棲息地。
基因技術在蛙類保育中的應用是另一個快速發展的領域。隨著基因組測序技術的進步和成本的降低,科學家們能夠更深入地了解蛙類的遺傳多樣性和適應能力。這些信息對於制定有效的保育策略至關重要。
一個重要的應用是通過基因分析來評估種群的健康狀況和遺傳多樣性。例如,在美國加州,研究人員使用基因標記來研究紅腿蛙(Rana draytonii)的種群結構和基因流動。這些信息幫助保育者確定哪些種群需要優先保護,以及如何設計廊道來促進種群間的基因交流。
基因編輯技術,如CRISPR-Cas9,也在蛙類保育中展現出潛力。雖然這項技術的應用仍處於早期階段,但科學家們正在探索如何利用它來增強蛙類對疾病的抵抗力。例如,有研究團隊正在試圖識別和強化能夠抵抗壺菌的基因。然而,將基因編輯技術應用於野生種群仍然存在倫理和生態風險,需要謹慎評估。
另一個基因技術的應用是環境DNA(eDNA)監測。通過分析水體或土壤中的DNA痕跡,科學家們可以檢測到蛙類的存在,而無需直接觀察或捕捉它們。這種非侵入性的方法特別適用於稀有或難以發現的物種。例如,在法國的一項研究中,研究人員成功使用eDNA技術來監測濕地中的多個蛙類物種,包括一些瀕危物種。
基因庫的建立也是基因技術在蛙類保育中的重要應用。通過保存瀕危蛙類的遺傳材料,科學家們可以為未來的保育工作提供寶貴的資源。例如,"冰凍方舟"項目正在收集和冷凍保存世界各地瀕危兩棲動物的精子和卵子。這些材料可能在未來用於重建已經滅絕的種群。
公民科學在蛙類研究與保育中正在發揮越來越重要的作用。通過邀請公眾參與數據收集和監測工作,科學家們可以獲得大規模、長期的數據,這些數據對於了解蛙類分布和種群動態至關重要。
一個成功的例子是全球範圍內的"蛙鳴監測計劃"。這個項目邀請志願者們記錄他們所在地區的蛙鳴聲。參與者可以使用智能手機應用程序來識別和記錄蛙類的叫聲。這些數據不僅幫助科學家追踪蛙類的分布變化,還能監測繁殖季節的變化,這對於了解氣候變化的影響特別有價值。
另一個公民科學項目是"失蹤的蛙類"計劃。這個全球性的項目鼓勵公眾尋找已經多年未被發現的蛙類物種。通過這個項目,一些被認為已經滅絕的物種被重新發現,如在厄瓜多爾重新發現的棱牆樹蛙(Hyloscirtus pantostictus)。
公民科學還在蛙類棲息地保護中發揮重要作用。例如,在澳大利亞,"蛙類友好型後院"項目鼓勵居民創造適合蛙類生存的花園環境。參與者學習如何建造小型池塘、選擇合適的植物,並避免使用有害的化學品。這不僅直接施惠蛙類,還提高了公眾對蛙類保護的意識。
公民科學項目還有助於監測和控制入侵物種。例如,在佛羅里達州,志願者們參與了對海地蟾蜍(Rhinella marina)的監測和移除工作。這種入侵物種對本地生態系統造成了嚴重威脅,公民科學家的參與大大提高了控制計劃的效率。
然而,公民科學也面臨一些挑戰,如數據質量控制和參與者培訓等。為此,許多項目開發了培訓材料和數據驗證系統,以確保收集的信息的準確性和可靠性。
蛙類保育的未來挑戰與機會是多方面的,需要綜合考慮生態、社會和經濟因素。
首先,氣候變化無疑是蛙類面臨的最大挑戰之一。氣候變化不僅直接影響蛙類的棲息地,還可能改變食物鏈、加劇疾病傳播,並與其他威脅因素產生協同效應。應對這一挑戰需要多層面的策略,包括減緩氣候變化的全球努力,以及幫助蛙類適應變化的局部措施。例如,設計氣候智能型保護區網絡,確保在未來氣候條件下仍然有適合的棲息地。
其次,棲息地破碎化和喪失仍然是許多蛙類面臨的主要威脅。隨著人類活動的擴張,蛙類的生存空間不斷縮小。未來的保育工作需要更好地整合土地使用規劃和生態保護。一個可能的方向是發展"綠色基礎設施",如生態廊道和都市濕地,以連接分散的棲息地。此外,恢復退化的生態系統也將是一個重要的機會。
疾病管理將持續成為蛙類保育的一個關鍵挑戰。除了已知的威脅如壺菌病,新的病原體可能會隨著全球氣候變化和人類活動的擴張而出現。這要求我們加強疾病監測系統,開發新的治療和預防方法。基因技術和微生物學的進步可能為此提供新的工具。
污染,尤其是新興污染物如微塑料和藥物殘留,對蛙類的影響還有待深入研究。這些污染物可能以複雜的方式影響蛙類的生理和行為,研究它們的長期影響將是未來的一個重要方向。
入侵物種的管理也將持續是一個挑戰。隨著全球貿易和旅行的增加,更多的外來物種可能被引入到新的環境中。開發有效和人道的控制方法,同時預防新的入侵,將是未來工作的重點。
在機遇方面,技術進步為蛙類研究和保育帶來了新的可能性。例如,人工智能和機器學習技術可能徹底改變蛙類監測和數據分析。無人機技術可以幫助我們更好地調查和監測難以到達的棲息地。基因組學技術的進步可能為我們提供更多工具來了解和保護蛙類的遺傳多樣性。
另一個重要的機遇是跨學科合作的加強。蛙類保育不僅需要生物學家和生態學家,還需要氣候科學家、社會科學家、經濟學家和政策制定者的參與。例如,將生態系統服務的概念納入經濟決策中,可以為蛙類棲息地的保護提供更強的動力。
教育和公眾參與也提供了重要的機會。隨著環境意識的提高,越來越多的人願意參與到保育工作中來。發展創新的教育計劃,利用新媒體技術來傳播保育信息,將有助於建立更廣泛的社會支持。
國際合作的加強也是一個重要機遇。許多蛙類的棲息地跨越國界,有效的保育需要國際協調。加強數據共享、制定共同的保育策略,以及協調跨境保護區的管理,都可能帶來積極的影響。
最後,將蛙類保育與更廣泛的可持續發展目標相結合是未來的一個重要機遇。蛙類保育不應被視為孤立的工作,而應該與水資源管理、氣候變化適應、生物多樣性保護等更大的環境目標相結合。這種整合方法不僅可以為蛙類保育爭取更多資源和支持,還可以產生多重效益。
例如,在城市規劃中納入蛙類友好的設計,不僅可以為蛙類提供棲息地,還可以改善城市的水資源管理和微氣候。在荷蘭的鹿特丹,一個創新的項目正在將屋頂花園、雨水收集系統和兩棲動物棲息地結合起來,這不僅為城市蛙類提供了生存空間,還幫助減少了城市洪水風險和熱島效應。
在農業領域,推廣對蛙類友好的耕作方式可以與可持續農業實踐相結合。例如,在水稻種植中保留部分濕地區域不僅可以為蛙類提供棲息地,還可以幫助控制害蟲,減少農藥使用。在日本的某些地區,農民正在實施這種"蛙類友好型"水稻種植方法,不僅保護了當地的蛙類種群,還生產出了高品質的有機大米,為農民帶來了額外的經濟效益。
在未來,基於自然的解決方案(Nature-based Solutions)可能成為蛙類保育的一個重要方向。這種方法強調利用自然生態系統來應對環境挑戰,同時保護生物多樣性。例如,利用濕地來淨化水質和控制洪水,這不僅可以為蛙類提供棲息地,還能為人類社區帶來多重效益。在中國,長江流域的一些地區正在實施"海綿城市"計劃,通過恢復和建造濕地來管理城市水資源,這同時也為蛙類等兩棲動物創造了棲息環境。
另一個值得關注的未來方向是將蛙類保育與生態旅遊結合起來。通過設計適當的生態旅遊項目,不僅可以為保育工作籌集資金,還可以提高公眾對蛙類和濕地生態系統的認識。例如,在哥斯達黎加的蒙特維德雲霧森林,針對樹蛙的生態旅遊項目不僅成功地保護了當地的蛙類種群,還為當地社區帶來了經濟效益。這種模式可能在其他地區得到推廣,特別是在生物多樣性豐富但經濟相對落後的地區。
在研究方面,跨學科和跨領域的合作將變得越來越重要。例如,將生態學研究與社會科學結合起來,可以幫助我們更好地理解人類行為對蛙類的影響,以及如何設計更有效的保育政策。在巴西的亞馬遜地區,一個研究項目正在結合生態學、人類學和經濟學的方法,研究當地社區的土地使用方式如何影響蛙類多樣性,並探索可持續的土地管理模式。
此外,新興的技術如環境DNA(eDNA)分析、遠程感應和人工智能等,將為蛙類研究和監測提供新的工具。這些技術可以幫助我們更有效地檢測稀有物種,監測種群變化,並預測未來的分布模式。例如,在加拿大的不列顛哥倫比亞省,研究人員正在使用eDNA技術來監測瀕危的斑點蛙(Rana luteiventris)的分布,這種方法比傳統的實地調查更加高效和低干擾。
基因技術的應用也可能在未來的蛙類保育中發揮更大的作用。除了前面提到的應用外,基因編輯技術可能被用來增強瀕危物種的適應能力。例如,通過基因編輯來增強蛙類對特定疾病的抵抗力。然而,這種方法也帶來了倫理和生態風險,需要謹慎評估和管理。
在面對氣候變化的挑戰時,適應性管理將成為蛙類保育的一個關鍵策略。這意味著保育計劃需要更加靈活,能夠根據不斷變化的環境條件和新的科學發現進行調整。例如,設計動態的保護區網絡,能夠隨著氣候變化而調整邊界和管理策略。在澳大利亞的大堡礁地區,科學家們正在開發這樣的適應性管理模型,以應對氣候變化對當地蛙類和其他野生動物的影響。
城市化繼續快速發展,城市生態學在蛙類保育中的重要性將會增加。未來的研究可能會更多地關注如何在城市環境中創造和維護適合蛙類生存的棲息地。這可能包括設計多功能的城市綠地,將蛙類棲息地與雨水管理和休閒空間結合起來。在新加坡,"花園城市"的理念正在被擴展到包括為蛙類等野生動物創造棲息地,例如在公園和住宅區中建造生態池塘。
在全球範圍內,跨境合作將成為蛙類保育的一個重要趨勢。許多蛙類的棲息地跨越國界,有效的保育需要國際協調。這可能包括建立跨境保護區、協調保育政策、共享數據和資源等。例如,在湄公河流域,中國、老撾、泰國和越南正在合作保護共享的濕地生態系統,這對於該地區的蛙類保育至關重要。
公民科學在蛙類研究和保育中的作用可能會進一步擴大。隨著智能手機和其他便攜設備的普及,公眾參與數據收集的能力大大增強。未來可能會看到更多創新的公民科學項目,如利用人工智能輔助的蛙類叫聲識別應用程序,或者是基於遊戲化設計的蛙類監測平台。這不僅可以收集大量有價值的數據,還能提高公眾的環境意識。
在教育方面,虛擬現實(VR)和增強現實(AR)技術可能為蛙類保育教育帶來新的可能性。這些技術可以讓人們身臨其境地體驗蛙類的生活環境,了解它們面臨的威脅,從而增強保育意識。例如,一些自然歷史博物館正在開發VR項目,讓訪客能夠"縮小"到蛙類的大小,體驗它們的世界。
隨著基因組學技術的進步,我們對蛙類適應性和進化的理解將會深化。這可能導致更精確的保育策略,例如基於基因組信息的種群管理計劃。同時,這也可能帶來新的挑戰,如如何在保護遺傳多樣性的同時促進適應性進化。
在應對新興疾病威脅方面,預警系統和快速反應機制的建立將變得越來越重要。這可能包括全球範圍內的蛙類健康監測網絡,以及開發新的疾病診斷和治療方法。例如,一些研究團隊正在探索使用益生菌來增強蛙類對疾病的抵抗力。
最後,將蛙類保育納入更廣泛的生態系統服務評估和自然資本核算中,可能成為未來的一個重要趨勢。這種方法可以幫助我們更好地量化蛙類及其棲息地的經濟價值,為保育工作提供更強有力的政策支持。例如,在評估濕地的生態系統服務時,考慮蛙類在害蟲控制、養分循環等方面的貢獻。
總的來說,蛙類研究與保育的未來方向將更加綜合、跨學科,並更緊密地與全球環境挑戰和可持續發展目標相結合。這需要研究者、保育者、政策制定者和公眾的共同努力,以確保這些獨特而重要的生物在不斷變化的世界中繼續繁衍生息。
在蛙類研究與保育的未來方向中,我們還需要考慮一些新興的領域和方法。
微生物組研究可能成為蛙類保育的一個重要新方向。蛙類皮膚上的微生物群落對其健康和疾病抵抗力有重要影響。未來的研究可能會更深入地探索如何利用有益微生物來增強蛙類的疾病抵抗力,特別是對抗如壺菌病等致命疾病。例如,在美國的黃石國家公園,科學家們正在研究某些蛙類種群如何通過皮膚微生物組來抵抗壺菌感染,希望能夠開發出新的保育策略。
此外,環境污染物對蛙類的影響也需要更多關注。新興污染物如微塑料、藥物殘留和內分泌干擾物對蛙類的長期影響尚不清楚。未來的研究可能會更多地關注這些污染物如何影響蛙類的生理、行為和種群動態。在瑞典,研究人員正在調查農業地區的殺蟲劑殘留如何影響當地蛙類的內分泌系統和繁殖能力,這類研究對於制定更有效的環境政策至關重要。
在保育策略方面,"共生設計"(Symbiotic Design)可能成為一個創新方向。這種方法旨在創造既滿足人類需求又有利於野生動物的環境。例如,在城市規劃中,可以設計既能管理雨水又能為蛙類提供棲息地的多功能綠地。在荷蘭的阿姆斯特丹,一個創新項目正在將屋頂花園、雨水收集系統和兩棲動物棲息地結合起來,這不僅為城市蛙類提供了生存空間,還幫助減少了城市洪水風險。
人工智能(AI)和大數據分析在蛙類研究中的應用可能會進一步擴大。這些技術可以幫助我們更有效地分析大量的生態數據,預測蛙類分布變化,並優化保育策略。例如,機器學習算法可以用來分析衛星圖像,自動識別和監測適合蛙類生存的棲息地。在巴西的亞馬遜地區,研究人員正在使用AI技術來分析大量的聲音記錄,以監測蛙類多樣性的變化。
在氣候變化背景下,"輔助遷移"(Assisted Migration)可能成為一個具有爭議但潛在必要的保育策略。這涉及將蛙類物種人為遷移到氣候變化下可能更適合的新棲息地。雖然這種方法存在生態風險,但對於某些面臨滅絕威脅的物種可能是必要的。在澳大利亞,科學家們正在評估是否需要將一些高山蛙類遷移到更高海拔的地區,以應對氣候變暖。
社會生態系統方法在蛙類保育中的應用可能會增加。這種方法認識到人類社會和生態系統是密切相連的,強調在保育策略中考慮社會、文化和經濟因素。例如,在設計保護區時,不僅要考慮生態因素,還要考慮當地社區的需求和傳統使用權。在哥斯達黎加,一些保護區正在採用這種方法,將蛙類保育與社區發展和生態旅遊相結合。
在基因保育方面,基因庫和冷凍保存技術可能會得到更廣泛的應用。這些技術可以保存瀕危蛙類的遺傳材料,為未來的種群恢復提供可能性。例如,在澳大利亞,研究人員正在為多種瀕危蛙類建立精子和胚胎庫,作為對抗可能的滅絕的後備計劃。
跨學科研究在蛙類保育中的重要性將進一步增加。例如,將生態學、流行病學和氣候科學結合起來,可以幫助我們更好地理解和預測疾病對蛙類的影響。在美國加州,一個跨學科團隊正在研究氣候變化、疾病傳播和蛙類種群動態之間的複雜關係。
在公眾參與方面,"公民科學家"的角色可能會進一步擴大和專業化。未來可能會看到更多專門培訓的志願者參與到複雜的監測和研究項目中。例如,在英國,一些保育組織正在培訓志願者進行專業的蛙類健康檢查和DNA採樣。
生態系統服務支付(Payment for Ecosystem Services)機制可能會在蛙類棲息地保護中發揮更大作用。例如,為保護重要的蛙類棲息地,可以向土地所有者提供經濟補償。在哥斯達黎加,已經有類似的計劃在運作,農民因保護森林和濕地而獲得補償,這間接施惠了當地的蛙類種群。
在教育和公眾參與方面,虛擬和增強現實技術可能會被更廣泛地應用於蛙類保育教育。這些技術可以創造身臨其境的體驗,讓人們更好地理解蛙類的生活環境和面臨的威脅。例如,一些自然歷史博物館正在開發VR項目,讓訪客能夠"變身"成蛙類,體驗它們的世界。
最後,隨著環境DNA(eDNA)技術的發展,我們可能會看到更多的大規模、低干擾性的蛙類監測項目。這種方法可以通過分析水樣中的DNA來檢測蛙類的存在,特別適用於稀有或難以直接觀察的物種。在加拿大的不列顛哥倫比亞省,研究人員正在使用eDNA技術來監測大範圍內的蛙類分布情況。
這些新興的研究方向和保育策略反映了蛙類保育工作的複雜性和多樣性。它們強調了跨學科合作的重要性,以及將蛙類保育與更廣泛的環境和社會問題相結合的必要性。隨著這些新方法和技術的發展,我們有望更有效地保護這些重要的兩棲動物,確保它們在不斷變化的世界中繼續繁衍生息。