1. 蜘蛛世界簡介
蜘蛛是一種令人著迷的生物,牠們在地球上已經存在了數億年。蜘蛛屬於節肢動物門的蛛形綱,與昆蟲有著密切的親緣關係,但在演化過程中發展出了獨特的特徵。全球已知的蜘蛛種類超過48,000種,分布在各種生態系統中,從熱帶雨林到極地苔原,甚至是都市環境中都能找到牠們的蹤跡。
蜘蛛在生態系統中扮演著重要的角色,主要作為捕食者控制其他節肢動物的數量。牠們的存在有助於維持生態平衡,特別是在農業生態系統中,蜘蛛可以有效地控制害蟲數量,減少農藥的使用。此外,蜘蛛的蛛絲是自然界中最堅韌的材料之一,具有極高的應用價值,科學家正在研究如何將其應用於醫療、紡織和工程等領域。
在人類文化中,蜘蛛常常被賦予豐富的象徵意義。在許多古老的神話和傳說中,蜘蛛被視為智慧、創造力和耐心的化身。例如,在希臘神話中,織女阿拉克涅(Arachne)因挑戰女神雅典娜(Athena)的織藝而被變成蜘蛛。在某些非洲和美洲原住民文化中,蜘蛛被視為宇宙的創造者或是帶來好運的象徵。
然而,對於許多人來說,蜘蛛也是恐懼和迷思的來源。常見的誤解包括認為所有蜘蛛都有劇毒、蜘蛛會主動攻擊人類等。事實上,絕大多數蜘蛛對人類無害,只有極少數種類的蜘蛛咬傷可能對人體造成嚴重影響。因此,了解蜘蛛的真實特性,消除不必要的恐懼,對於促進人類與這些重要生物和諧共處至關重要。
1.1 什麼是蜘蛛?
蜘蛛是一類獨特而引人入勝的節肢動物,屬於蛛形綱(Arachnida)中的蜘蛛目(Araneae)。這些生物以其多樣性和適應能力而聞名,在地球上幾乎所有的陸地生態系統中都能找到牠們的蹤跡。蜘蛛與昆蟲有著密切的親緣關係,但在演化過程中發展出了許多特有的特徵,使其成為一個獨特的類群。
蜘蛛的身體結構是其最顯著的特徵之一。牠們的身體分為兩個主要部分:前體部(prosoma)和後體部(opisthosoma)。前體部又稱為頭胸部,包含了蜘蛛的眼睛、口器、毒腺和腿部。大多數蜘蛛擁有八隻腿,這是區別牠們與昆蟲的一個重要特徵。蜘蛛的眼睛通常有八隻,arranged成不同的模式,但有些洞穴物種可能完全沒有眼睛。後體部則包含了蜘蛛的內部器官,如心臟、消化系統、生殖系統,以及最為人知的絲腺和絲疣。
蜘蛛的絲腺和絲疣是牠們最為獨特的器官之一。這些特化的腺體能夠產生蛛絲,這是一種極其堅韌且富有彈性的蛋白質纖維。蜘蛛利用這些絲來建造網、包裹獵物、製作蛋囊、以及作為「安全繩」在空中移動。每種蜘蛛都能產生多種不同類型的絲,每種絲都有其特定的用途。例如,有些絲用於構建網的框架,而另一些則用於捕獲獵物。
蜘蛛的捕食方式也是牠們的一大特色。絕大多數蜘蛛都是掠食性動物,主要以其他節肢動物為食。牠們採用多種捕獵策略,包括織網捕獲、主動追捕、以及伏擊等方式。蜘蛛通過注射毒液來制伏獵物,這種毒液不僅能麻痺獵物,還能開始消化過程,使蜘蛛能夠吸食獵物的體液。
在感覺系統方面,蜘蛛展現出令人驚嘆的適應性。除了視覺之外,牠們還高度依賴振動感應。蜘蛛的腿上分布著大量的感覺毛,能夠檢測空氣和表面的細微振動。這使得蜘蛛能夠精確地感知周圍環境的變化,無論是獵物的存在還是潛在威脅的接近。
蜘蛛的繁殖行為也十分有趣。雄性蜘蛛通常比雌性小,在求偶過程中常常表現出複雜的行為模式。有些種類的雄蛛會進行精心策劃的求偶舞蹈,或是給雌蛛帶來「求偶禮物」。然而,在某些物種中,雌蛛可能會在交配後或甚至交配過程中吃掉雄蛛,這種現象被稱為性食同類(sexual cannibalism)。
蜘蛛的生活史也值得關注。大多數蜘蛛從蛋中孵化,經過幼蛛階段,然後通過多次蛻皮成長為成蛛。有些蜘蛛種類的壽命僅有幾個月,而另一些則可以活幾年甚至幾十年。例如,某些塵蜘蛛(trapdoor spiders)的雌性個體據報道可以活到40歲以上。
在生態系統中,蜘蛛扮演著至關重要的角色。作為廣泛分布的捕食者,牠們對控制其他節肢動物的數量起著關鍵作用。在農業生態系統中,蜘蛛是重要的生物防治力量,能夠有效控制多種農業害蟲。此外,蜘蛛還是許多鳥類、爬行動物和小型哺乳動物的食物來源,在食物網中佔據重要地位。
蜘蛛的多樣性是驚人的。從體型微小如針尖的矮蛛(Patu digua),到腿展可達30厘米的鳥蛛(Theraphosa blondi),蜘蛛的大小範圍極為廣泛。牠們的外形和行為也千變萬化,有些蜘蛛能夠完美地偽裝成樹皮或花朵,而另一些則以鮮艷的色彩警告潛在捕食者。有些蜘蛛生活在水下,建造可以儲存空氣的潛水鐘;還有些蜘蛛能夠在水面上「行走」,甚至捕食小魚。
人類對蜘蛛的認知和態度一直在變化。在許多文化中,蜘蛛被視為智慧和創造力的象徵。然而,對蜘蛛的恐懼(蜘蛛恐懼症)在人群中也相當普遍。這種恐懼往往源於對蜘蛛的誤解,如認為所有蜘蛛都具有致命的毒性。事實上,儘管大多數蜘蛛確實有毒,但只有極少數物種的毒液對人類有真正的威脅。
隨著科學研究的深入,人們對蜘蛛的了解不斷增加。蜘蛛不僅在生態學研究中扮演重要角色,在生物技術領域也展現出巨大潛力。例如,科學家們正在研究如何利用蛛絲的獨特properties來開發新型材料,這些材料可能應用於醫療、航空航太等多個領域。此外,蜘蛛毒液中的某些成分也被發現具有潛在的醫療價值,可能用於開發新型藥物。
1.2 蜘蛛在生態系統中的重要性
蜘蛛在生態系統中扮演著舉足輕重的角色,其重要性遠超過大多數人的認知。這些八腳動物不僅是自然界中最成功的捕食者之一,還是維持生態平衡的關鍵物種。蜘蛛的存在對於控制其他節肢動物的數量、促進生物多樣性、以及支持整個食物網的運作都有著深遠的影響。
首先,蜘蛛是極其高效的捕食者。牠們主要以其他節肢動物為食,包括許多被視為害蟲的昆蟲。在農業生態系統中,蜘蛛的這一特性尤為重要。研究表明,在一公頃的農田中,蜘蛛每年可以捕食多達數百公斤的昆蟲。這種自然的害蟲控制機制大大減少了農民對化學農藥的依賴,從而降低了環境污染和生產成本。例如,在中國的水稻田中,研究人員發現增加蜘蛛的數量可以顯著降低稻飛蝨等害蟲的危害,提高作物產量。
蜘蛛的捕食行為不僅限於農田,在森林、草原、沙漠等各類生態系統中,牠們都發揮著類似的作用。通過控制昆蟲和其他小型節肢動物的數量,蜘蛛幫助維持這些生態系統的平衡。例如,在北美的森林中,蜘蛛是控制森林害蟲松毛蟲數量的重要因素之一。當松毛蟲爆發時,蜘蛛的數量也會相應增加,從而幫助遏制害蟲的蔓延。
蜘蛛的存在還有助於促進生物多樣性。作為中層捕食者,蜘蛛不僅控制獵物的數量,也為其他動物提供食物來源。許多鳥類、爬行動物、兩棲動物和小型哺乳動物都以蜘蛛為食。例如,北美的蜂鳥在築巢期間大量捕食蜘蛛,以獲取豐富的蛋白質來餵養幼鳥。這種複雜的食物網關係有助於維持生態系統的穩定性和多樣性。
在水生生態系統中,某些特化的蜘蛛種類也發揮著重要作用。例如,漁蛛(Dolomedes)能夠在水面上行走,捕食水生昆蟲和小魚。這些蜘蛛成為了連接水陸生態系統的重要紐帶,參與了能量和物質在不同生態系統間的流動。
蜘蛛還通過其獨特的織網行為改變局部環境,創造了微生境。蛛網不僅是捕獵的工具,還為其他小型生物提供了庇護所。一些小型昆蟲和微生物會利用廢棄的蛛網作為棲息地,形成了以蛛網為中心的微型生態系統。這種「生態系統工程師」的角色進一步增加了環境的複雜性和生物多樣性。
在生物指標方面,蜘蛛也顯示出了重要價值。由於蜘蛛對環境變化敏感,科學家們常常使用蜘蛛群落的組成和多樣性來評估生態系統的健康狀況。例如,在歐洲的一些研究中,科學家通過監測特定生境中蜘蛛種類的變化來評估氣候變化對生態系統的影響。這為生態系統管理和保護提供了寶貴的信息。
蜘蛛在養分循環中也扮演著重要角色。當蜘蛛捕食昆蟲後,其排泄物和未被完全消化的獵物殘骸會落入土壤,成為重要的有機養分來源。這些物質被分解者進一步分解,最終被植物吸收利用。在一些貧瘠的生態系統中,如沙漠或高山地區,蜘蛛的這一作用尤為重要,有助於維持脆弱生態系統的養分平衡。
蜘蛛的蛛絲在生態系統中也發揮著多重作用。除了用於捕獵,蛛絲還被許多鳥類用作築巢材料。一些研究發現,使用蛛絲築巢的鳥類,其巢穴更為堅固,幼鳥的存活率更高。此外,一些蜘蛛物種的幼體會利用蛛絲進行「氣球飛行」,這種行為有助於蜘蛛的長距離擴散,對維持種群基因交流和拓展新棲息地至關重要。
在城市生態系統中,蜘蛛的角色同樣不可忽視。隨著城市化進程的加快,許多蜘蛛物種已經適應了城市環境。這些「城市蜘蛛」在控制蚊子、蒼蠅等城市害蟲方面發揮著重要作用,為城市居民提供了免費的生物防治服務。例如,在澳大利亞的一項研究中,科學家發現城市花園中的蜘蛛每年可以捕食相當於整個社區居民體重的昆蟲。
蜘蛛還通過其獨特的生理特性為人類提供了寶貴的研究素材。蜘蛛毒液中含有大量複雜的生物活性化合物,其中一些已被證明具有潛在的醫療價值。例如,從智利烏蜘蛛(Latrodectus mactans)毒液中提取的一種化合物被發現可能有助於治療勃起功能障礙。此外,蜘蛛的蛛絲因其獨特的力學性質,正被科學家們研究用於開發新型材料,這些材料可能在醫療、航空航太等領域有廣泛應用。
蜘蛛在生態系統中的重要性還體現在其作為進化研究的模型生物。由於蜘蛛種類繁多,適應性強,科學家們可以通過研究不同蜘蛛種類的形態、行為和生理特性來探索生物如何適應不同環境。這些研究不僅有助於我們理解進化過程,還為保護生物多樣性提供了重要依據。
1.3 蜘蛛與人類文化
蜘蛛在人類文化中佔有獨特而複雜的地位,從遠古時代起就在神話、傳說、藝術和文學中扮演著重要角色。這些八腳生物在不同文化中往往具有多層面的象徵意義,既被視為智慧和創造力的化身,又常常引發恐懼和敬畏。蜘蛛與人類文化的緊密聯繫反映了我們與這些神秘生物之間長期而複雜的關係。
在許多古老文化中,蜘蛛被賦予了創造者的身份。例如,在西非的阿肯(Akan)文化中,蜘蛛神安南西(Anansi)被視為智慧和故事的守護者。安南西常常以狡猾的騙子形象出現在民間故事中,通過機智和詭計戰勝比自己強大的對手。這些故事不僅娛樂了人們,還傳遞了重要的文化價值觀和生存智慧。類似地,在北美的納瓦霍(Navajo)族文化中,蜘蛛女神被認為教導了人類織布的技藝,象徵著創造和智慧。
在古希臘神話中,蜘蛛與織布藝術密切相關。最著名的故事是關於織女阿拉克涅(Arachne)的傳說。阿拉克涅是一位技藝精湛的織女,她挑戰女神雅典娜(Athena)比試織布。雖然阿拉克涅的作品無可挑剔,但她的傲慢激怒了雅典娜。作為懲罰,阿拉克涅被變成了蜘蛛,注定要永遠織網。這個神話不僅解釋了蜘蛛的由來,還反映了古希臘人對技藝和謙遜的看法。
在中國文化中,蜘蛛通常被視為吉祥的象徵。「蜘蛛」的諧音「知住」,意味著好運將會降臨。因此,在中國的民間傳說中,看到蜘蛛從天而降被認為是好運即將到來的徵兆。這種正面的文化聯想與西方文化中常見的蜘蛛恐懼形成了鮮明對比。
在歐洲中世紀的基督教文化中,蜘蛛有時被賦予宗教意義。例如,有一個廣為流傳的傳說講述了聖母瑪利亞和耶穌逃往埃及時,一隻蜘蛛在洞口織網,掩護了他們免受追兵發現。這個故事將蜘蛛塑造成了上帝的助手,強調了看似微不足道的生物也能在神聖計劃中發揮重要作用。
然而,在許多文化中,蜘蛛也常常與恐懼和危險聯繫在一起。這種恐懼可能源於蜘蛛的外形和行為,以及對其毒性的誇大認知。在歐洲民間傳說中,蜘蛛有時被視為女巫的化身或惡魔的使者。這種負面形象在中世紀的藝術和文學中得到了廣泛反映,並持續影響著現代文化。
蜘蛛在文學作品中也經常出現,往往帶有深刻的象徵意義。例如,在夏洛特·勃朗特(Charlotte Bronte)的小說《夏洛特的網》中,蜘蛛夏洛特成為了友誼、犧牲和創造力的象徵。她通過在網上織出單詞來拯救小豬威爾伯的生命,展現了語言的力量和創造性思維的重要性。
在現代流行文化中,蜘蛛的形象得到了進一步發展和利用。漫威漫畫中的超級英雄蜘蛛人(Spider-Man)就是一個典型例子。彼得·帕克因被放射性蜘蛛咬傷而獲得超能力,這個設定既反映了人們對蜘蛛力量的敬畏,也體現了現代科技時代人們對基因變異的想像。蜘蛛人的故事不僅娛樂了全球數億觀眾,還傳遞了「能力越大,責任越大」的道德理念。
在視覺藝術領域,蜘蛛也是一個受歡迎的主題。法國雕塑家路易絲·布爾喬亞(Louise Bourgeois)創作的巨型蜘蛛雕塑「媽媽」(Maman)就是一個著名例子。這些高達9米的青銅蜘蛛雕塑被安置在世界各地的著名美術館和公共空間中,引發了觀眾對母性、保護和創造力的深入思考。
在一些原住民文化中,蜘蛛在宗教儀式和傳統醫療中扮演著重要角色。例如,在南美的一些土著部落中,人們相信某些蜘蛛種類具有治療能力。他們會將特定種類的蜘蛛磨成粉末,用於治療各種疾病。這種做法雖然缺乏科學依據,但反映了人類尋求利用自然資源治療疾病的長期努力。
蜘蛛在現代科技和產品設計中也找到了自己的位置。例如,汽車製造商雷克薩斯(Lexus)的設計師曾從蜘蛛的八隻腿中獲得靈感,設計出了一款概念車的懸掛系統。這種設計靈感的跨界應用展示了自然界生物特徵如何啟發人類的技術創新。
在環境教育和科普活動中,蜘蛛常常被用作激發公眾對自然世界興趣的切入點。許多自然歷史博物館和動物園都設有蜘蛛展區,通過互動展示和教育活動,幫助人們克服對蜘蛛的恐懼,了解這些生物的生態重要性。這些努力在改變公眾對蜘蛛的認知、促進生物多樣性保護方面發揮了重要作用。
蜘蛛在人類文化中的地位是動態變化的,反映了社會價值觀和科學認知的演變。隨著人們對蜘蛛生態作用的深入了解,以及環境保護意識的提高,蜘蛛在現代文化中的形象正在逐漸改善。越來越多的人開始欣賞蜘蛛的美麗和生態價值,而不是單純地將其視為可怕或有害的生物。
1.4 常見的蜘蛛迷思
蜘蛛作為自然界中最常見且最具神秘色彩的生物之一,長期以來一直是各種迷思和誤解的主角。這些迷思不僅影響了人們對蜘蛛的認知,還可能導致不必要的恐懼和錯誤的行為。因此,理解和澄清這些常見的蜘蛛迷思對於促進人類與這些重要生物的和諧共處至關重要。
其中最廣為流傳的迷思之一是「所有蜘蛛都有劇毒」。事實上,雖然幾乎所有蜘蛛都能產生毒液,但絕大多數蜘蛛的毒性對人類而言是無害的。全球已知的約48,000種蜘蛛中,只有極少數種類的毒液對人體有實質性威脅。例如,在澳大利亞,雖然有許多毒蛛,但自1979年引入抗蜘蛛毒血清以來,還沒有因蜘蛛咬傷而導致的死亡案例。大多數蜘蛛的毒液主要用於麻痺和消化獵物,而不是對付體型遠大於它們的生物。
另一個常見的迷思是「蜘蛛會主動攻擊人類」。實際上,蜘蛛通常會盡量避開人類,只有在感到受到威脅時才會咬人。大多數蜘蛛咬人事件發生在人們不小心干擾或壓到蜘蛛時。蜘蛛的牙齒通常很小,許多種類甚至無法穿透人類的皮膚。因此,與其說蜘蛛是攻擊性動物,不如說它們是謹慎且易受驚嚇的生物。
「蜘蛛會在人睡覺時爬進嘴裡」這個迷思也廣為流傳。這個說法完全是杜撰的,沒有任何科學依據。蜘蛛對人類呼出的二氧化碳和身體散發的熱量非常敏感,會本能地遠離這些信號。此外,蜘蛛更喜歡安靜、陰暗的環境,人類的口腔顯然不符合這些條件。
許多人也相信「所有蜘蛛都會織網」。雖然織網確實是許多蜘蛛的特徵,但並非所有蜘蛛都會這樣做。例如,跳蛛(Salticidae)和狼蛛(Lycosidae)等蜘蛛不會織捕獵網,而是依靠敏捷的身手和銳利的視力來獵捕獵物。這些蜘蛛展示了蜘蛛種類的多樣性和適應性。
「蜘蛛是昆蟲」這個說法也是一個常見的誤解。蜘蛛雖然和昆蟲一樣屬於節肢動物門,但它們分屬不同的綱。蜘蛛屬於蛛形綱,而昆蟲屬於昆蟲綱。蜘蛛有八條腿、兩個體節,沒有觸角;而昆蟲有六條腿、三個體節,並且有一對觸角。這些差異反映了蜘蛛和昆蟲在演化過程中的分歧。
另一個流行的迷思是「蜘蛛每年要吃掉8隻蒼蠅才能生存」。這個說法源於一項誤解了的科學研究。實際上,蜘蛛的食物需求因種類和環境而異,有些蜘蛛可能幾週不進食也能存活,而有些則需要頻繁捕食。蜘蛛的飲食習慣遠比這個迷思描述的要複雜和多樣。
「蜘蛛網擁有未來科技材料的特性」這個說法雖然不完全是迷思,但常常被誇大。蛛絲確實是一種令人驚嘆的天然材料,具有優異的強度和彈性。然而,將其直接應用於高科技領域面臨諸多挑戰。科學家們正在努力研究如何模仿和改良蛛絲的特性,而不是直接使用蜘蛛產生的絲。
「所有蜘蛛都很小」這個觀點也是一個常見的誤解。雖然大多數蜘蛛確實體型較小,但也有一些巨型種類。例如,歌利亞鳥蛛(Theraphosa blondi)的腿展可達30厘米,體重可達170克。這種多樣性再次說明了蜘蛛適應不同生態位的能力。
「雄蛛總是比雌蛛小得多」這個說法雖然在許多蜘蛛種類中是正確的,但並不適用於所有蜘蛛。在某些種類中,雌雄蜘蛛的體型差異並不明顯。這種多樣性反映了不同種類蜘蛛在繁殖策略上的差異。
「蜘蛛可以預測天氣變化」這個說法部分正確,但常常被過度解讀。蜘蛛確實對環境變化敏感,可能會根據溫度、濕度等因素調整織網行為。然而,將這種行為直接等同於「預測天氣」是不準確的。蜘蛛的行為更多是對當前環境條件的反應,而不是對未來天氣的預測。
「所有蜘蛛都有八隻眼睛」這個說法也不完全正確。雖然大多數蜘蛛確實有八隻眼睛,但有些種類可能有六隻、四隻,甚至兩隻眼睛。一些生活在洞穴中的蜘蛛物種甚至完全沒有眼睛。這種變異反映了蜘蛛對不同生活環境的適應。
「蜘蛛會長出新的腿」這個說法部分正確,但需要澄清。蜘蛛確實有再生能力,但只限於未成年時期。成年蜘蛛失去的腿不會再生,但它們可以通過調整剩餘的腿來適應。
澄清這些迷思不僅有助於提高公眾對蜘蛛的認知,還能促進人們對這些重要生物的理解和欣賞。隨著科學研究的深入,我們對蜘蛛的了解不斷增加,許多曾經被視為理所當然的觀點正在被重新評估。這種認知的轉變不僅有助於消除不必要的恐懼,還能促進更好的生物多樣性保護策略。
2. 蜘蛛的演化與多樣性
蜘蛛的演化與多樣性是一個引人入勝的研究領域,揭示了這些八腳動物如何在地球上繁衍並適應各種環境。蜘蛛的起源可以追溯到大約4億年前的泥盆紀,最早的化石記錄顯示它們與現代蜘蛛已有顯著差異。隨著時間的推移,蜘蛛經歷了顯著的演化,發展出多種獨特的特徵,如絲腺、毒腺和特化的感官系統。
蜘蛛的多樣性令人驚嘆,目前已知的物種超過48,000種,分布在全球各種生態系統中。這種多樣性體現在形態、行為和生態適應等多個方面。例如,跳蛛(Salticidae)家族以其敏銳的視力和複雜的求偶舞蹈而聞名,而捲葉蛛(Araneidae)則以其精巧的圓形網而著稱。
蜘蛛的適應性輻射導致了各種獨特的生存策略。有些物種,如水蜘蛛(Argyroneta aquatica),已經適應了水下生活,能夠在水中建造充滿空氣的「潛水鐘」。另一些,如沙漠中的輪蛛(Carparachne aureoflava),則進化出了在沙丘上「翻滾」逃避捕食者的能力。
蜘蛛的多樣性不僅體現在形態上,還體現在行為和生態位的多樣性上。從獨居的狩獵者到具有複雜社會結構的群居物種,蜘蛛展示了廣泛的行為適應。這種多樣性使蜘蛛成為研究演化生物學、行為生態學和適應性進化的重要模型生物。
2.1 蜘蛛的起源
蜘蛛的起源是一個深邃而引人入勝的話題,涉及了地球生命演化的漫長歷程。這些八腳動物的祖先可以追溯到約4億年前的泥盆紀早期,當時地球上的生命正經歷著一個重要的轉折點,陸地生態系統開始蓬勃發展。
最早的蜘蛛化石記錄來自於德文郡系統(Devonian System)的地層中。這些化石揭示了原始蜘蛛與現代蜘蛛之間的顯著差異,同時也展示了一些關鍵的相似特徵。例如,在蘇格蘭萊恩河畔(Rhynie)發現的化石中,科學家們發現了一種名為帕萊奧蠅蛛(Palaeotarbus jerami)的生物,被認為是目前已知最古老的蜘蛛化石之一。這種生物雖然還不能完全確定是否為真正的蜘蛛,但它展示了許多蜘蛛的原始特徵。
蜘蛛被認為是從一種名為尤里翼蛛(Uraraneida)的古老節肢動物演化而來。尤里翼蛛是蜘蛛的直系祖先,它們生活在3.8億到2.9億年前。這些生物已經具備了一些蜘蛛的特徵,如產絲的能力,但它們還保留著一些更原始的特徵,如腹部末端的一對附肢,這在現代蜘蛛中已經消失。尤里翼蛛的發現為我們理解蜘蛛的演化過程提供了重要的線索。
在蜘蛛演化的早期階段,一個關鍵的創新是絲腺的發展。最初,蛛絲可能主要用於包裹卵或作為巢穴的襯裡。隨著時間的推移,蛛絲的用途逐漸擴大,演化出了用於捕獵、築巢和遷移的功能。這種多功能的絲腺系統成為了蜘蛛成功適應各種生態位的重要因素。
另一個重要的演化特徵是毒腺的發展。早期的蜘蛛可能沒有毒腺,而是依靠強壯的螯肢來捕獲和壓碎獵物。毒腺的出現大大提高了蜘蛛的捕獵效率,使它們能夠獵捕更大、更強壯的獵物。這一創新也為蜘蛛開拓了新的生態位,使它們能夠在食物鏈中佔據更重要的位置。
蜘蛛的感官系統也在演化過程中不斷完善。早期的蜘蛛可能主要依賴觸覺和振動感應來感知環境。隨著時間的推移,視覺系統逐漸發展,特別是在一些捕獵性強的蜘蛛科屬中,如跳蛛科(Salticidae)。這些蜘蛛發展出了複雜的眼睛結構,能夠精確地定位和追蹤獵物。
蜘蛛的形態學特徵在演化過程中也發生了顯著變化。早期的蜘蛛可能具有更分節的身體結構,類似於現代的鞭蠍。隨著時間的推移,蜘蛛的身體逐漸演化成為現在我們所熟知的兩體節結構:由頭胸部和腹部組成。這種身體結構的演變可能與蜘蛛的運動方式和捕獵策略的改變有關。
蜘蛛的繁殖策略也在演化過程中不斷完善。早期的蜘蛛可能採用較為簡單的繁殖方式,如直接將卵產在地面或水中。隨著時間的推移,蜘蛛演化出了複雜的求偶行為和精心設計的育兒策略。例如,一些蜘蛛物種發展出了精巧的求偶舞蹈,而另一些物種則進化出了攜帶卵囊或幼蛛的行為。
蜘蛛的生態適應能力在其演化歷程中扮演了關鍵角色。隨著陸地生態系統的複雜化,蜘蛛不斷適應新的環境和生態位。例如,一些蜘蛛物種適應了沙漠環境,發展出了特殊的行為和生理特徵來應對極端乾旱和高溫。另一些物種則適應了高山或極地環境,能夠在低溫和稀薄空氣中生存。
蜘蛛的社會性行為也是演化過程中的一個有趣方面。雖然大多數蜘蛛是獨居的,但有一些物種演化出了複雜的社會結構。例如,非洲的社會性蜘蛛安氏蛛(Stegodyphus dumicola)能夠形成大型的集體巢穴,共同捕獵和撫育後代。這種社會性行為的演化為我們理解動物社會的起源提供了寶貴的見解。
蜘蛛的起源和演化過程不僅反映了地球生命史的一個重要篇章,也為我們理解生物適應性和多樣性提供了豐富的案例。通過研究蜘蛛的演化歷程,科學家們得以深入了解生物如何應對環境變化、開拓新的生態位,以及在競爭激烈的生態系統中生存和繁衍。
蜘蛛的演化故事還在繼續,現代蜘蛛正面臨著新的環境挑戰,如氣候變化和棲息地破壞。研究蜘蛛的起源和演化不僅有助於我們理解過去,也為預測和保護這些重要生物的未來提供了重要依據。隨著新的化石發現和分子生物學技術的發展,我們對蜘蛛起源的認識也在不斷深化,揭示出更多令人驚嘆的演化奧秘。
2.2 主要的蜘蛛目和科
蜘蛛的分類體系反映了這些八腳動物驚人的多樣性和複雜的演化歷程。現代分類學將蜘蛛劃分為兩個主要亞目:原蛛亞目(Mesothelae)和新蛛亞目(Opisthothelae),後者又分為原腹蛛(Mygalomorphae)和真腹蛛(Araneomorphae)兩個支系。這種分類體系不僅反映了蜘蛛的演化關係,也體現了它們在形態和生態特徵上的顯著差異。
原蛛亞目是現存最原始的蜘蛛群體,僅包含一個科:隆蛛科(Liphistiidae)。這些蜘蛛保留了許多原始特徵,如腹部明顯分節和紡絲器位於腹部中部。隆蛛主要分布在東南亞地區,它們的存在為我們理解蜘蛛的早期演化提供了寶貴的線索。例如,日本的霧社隆蛛(Heptathela kikuyai)就是一種典型的原蛛,其形態特徵與化石記錄中的早期蜘蛛極為相似。
原腹蛛包括了許多大型和長壽的蜘蛛物種,如著名的鳥蛛科(Theraphosidae)。這個支系的蜘蛛通常具有粗壯的身體、強大的螯肢和相對簡單的絲腺系統。鳥蛛科中的歌利亞鳥蛛(Theraphosa blondi)是已知體型最大的蜘蛛,腿展可達30厘米。原腹蛛還包括了一些具有特殊生態適應的科,如澳洲的漏斗網蛛科(Atracidae),其中的雪梨漏斗網蛛(Atrax robustus)以其強力毒液而聞名。
真腹蛛是蜘蛛中最為多樣和普遍的群體,包含了絕大多數現代蜘蛛物種。這個支系的蜘蛛展現出了驚人的形態和行為多樣性。例如,園蛛科(Araneidae)的成員以其精巧的圓形網而著稱,如歐洲花園蜘蛛(Araneus diadematus)就是一個典型代表。跳蛛科(Salticidae)則以其敏銳的視力和複雜的求偶行為聞名,如孔雀跳蛛(Maratus volans)的雄性會展示色彩豔麗的腹部來吸引雌性。
狼蛛科(Lycosidae)的成員是活躍的地面獵手,它們不織網而是主動追捕獵物。狼蛛的一個顯著特徵是雌性會將卵囊附著在紡絲器上隨身攜帶,如歐洲狼蛛(Pardosa amentata)就展現了這種行為。球蛛科(Theridiidae)包括了許多常見的家居蜘蛛,如臭名昭著的黑寡婦蜘蛛(Latrodectus mactans),它以強力的神經毒素而聞名。
一些蜘蛛科展現出了獨特的生態適應。例如,昇蜘蛛科(Gnaphosidae)的成員主要在地面活動,它們的絲腺退化,反而發展出了強大的螯肢。蟹蛛科(Thomisidae)的蜘蛛則以其側行和擬態能力著稱,如花蟹蛛(Misumena vatia)能夠改變體色以匹配它們所棲息的花朵。
某些蜘蛛科展現出了令人驚嘆的特化適應。例如,盜蛛科(Mimetidae)的成員專門捕食其他蜘蛛,它們已經演化出了特殊的形態和行為來入侵其他蜘蛛的網並捕食網的主人。水蜘蛛科(Cybaeidae)中的水蜘蛛(Argyroneta aquatica)則完全適應了水下生活,能夠在水中建造充滿空氣的「潛水鐘」作為棲所。
一些蜘蛛科因其獨特的行為而引人注目。例如,梳蛛科(Deinopidae)中的網蛛(Deinopis spinosa)會織造一張特殊的網,當獵物接近時,蜘蛛會迅速將網拉伸並罩住獵物。社會性蜘蛛科(Eresidae)中的非洲社會性蜘蛛(Stegodyphus dumicola)則展現了罕見的群居行為,多個個體共同生活在大型的巢穴中。
某些蜘蛛科以其特殊的形態引人注目。例如,棘蛛科(Gasteracanthinae)的成員具有堅硬的外骨骼和棘狀突起,如六棘棘蛛(Gasteracantha cancriformis)就因其獨特的外形而聞名。長牙蛛科(Tetragnathidae)的成員則以其細長的身體和大型的螯肢為特徵,如金絲長牙蛛(Tetragnatha extensa)就是一個典型例子。
蜘蛛的分類體系不僅反映了它們的形態差異,也體現了它們的生態適應和行為特徵。例如,織網蜘蛛和非織網蜘蛛在分類上往往屬於不同的科,這反映了它們在捕獵策略上的根本差異。同樣,地棲蜘蛛和樹棲蜘蛛也常常屬於不同的分類群,反映了它們對不同棲息環境的適應。
隨著分子生物學技術的發展,蜘蛛的分類體系正在經歷持續的修訂和完善。新的DNA分析技術揭示了一些過去基於形態學難以發現的親緣關係,導致了一些分類位置的調整。例如,過去被歸類為單獨一科的毛蜘蛛(Ctenidae)現在被認為與狼蛛科有更密切的關係。
研究蜘蛛的主要目和科不僅有助於我們理解蜘蛛的多樣性,也為研究生物演化、生態適應和行為進化提供了豐富的素材。每一個蜘蛛科都代表了一個獨特的演化實驗,展示了生命如何應對各種環境挑戰並找到成功的生存策略。通過深入研究這些不同的蜘蛛群體,科學家們能夠更好地理解生物多樣性的形成過程和維持機制,為生物保護和生態管理提供重要的理論基礎。
2.3 蜘蛛的適應性輻射
蜘蛛的適應性輻射是演化生物學中一個引人入勝的主題,展示了這些八腳動物如何在地球上的各種生態位中成功擴散和多樣化。適應性輻射是指一個祖先物種在相對較短的時間內,因適應不同的環境而演化出多個後代物種的過程。蜘蛛在這方面展現出了驚人的能力,從原始的地棲捕食者演化成為了各種高度特化的形態和行為。
蜘蛛的適應性輻射可以從多個角度來觀察。首先是棲息地的多樣化。蜘蛛從最初的地面環境擴散到了幾乎所有的陸地生態系統,包括森林、草原、沙漠、高山,甚至是洞穴和水生環境。例如,在熱帶雨林中,我們可以看到樹冠層蜘蛛如黃金珠蛛(Nephila clavipes),它們織造巨大的捕獵網橫跨樹木之間。相比之下,在沙漠環境中,我們發現了如納米比亞輪蛛(Carparachne aureoflava)這樣的物種,它們能夠在沙丘上快速移動,甚至通過翻滾來逃避捕食者。
水生環境也見證了蜘蛛令人驚嘆的適應能力。歐洲水蜘蛛(Argyroneta aquatica)是唯一已知的完全水生蜘蛛,它們能夠在水下建造充滿空氣的「潛水鐘」作為棲所。這種適應展示了蜘蛛如何突破陸地生活的限制,成功進入水生生態位。
捕獵策略的多樣化是蜘蛛適應性輻射的另一個重要方面。雖然許多人將蜘蛛與織網聯繫在一起,但實際上蜘蛛展現了多種捕獵方式。織網蜘蛛如園蛛科(Araneidae)的成員,演化出了複雜的圓形網結構和行為,能夠有效捕捉飛行昆蟲。相比之下,跳蛛科(Salticidae)的成員則完全放棄了織網,而是依靠敏銳的視力和靈活的跳躍能力來追捕獵物。
一些蜘蛛物種的捕獵策略更加特化。例如,捲葉蛛(Phonognatha graeffei)會將一片捲起的樹葉整合到其網中作為隱蔽處,展示了環境利用的創新。另一個極端例子是南美洲的棒狀蜘蛛(Deinopis spinosa),它們會織造一種特殊的「投網」,當獵物接近時迅速將網拋出捕獲。
形態適應是蜘蛛適應性輻射的另一個顯著特徵。不同的生態位推動了蜘蛛形態的多樣化。例如,生活在樹皮上的蜘蛛如樹皮蟹蛛(Bassaniana versicolor)演化出扁平的體型,能夠緊貼樹皮表面。相比之下,生活在草叢中的蜘蛛如竹節蜘蛛(Tetragnatha)則發展出細長的體型,能夠在細小的植物莖上完美偽裝。
顏色和形態的擬態適應在蜘蛛中也很常見。例如,蟻形蜘蛛(Myrmarachne)不僅在外形上模仿螞蟻,還學會了模仿螞蟻的行為模式,這種複雜的擬態不僅幫助它們避免被捕食,還能夠接近和捕食真正的螞蟻。另一個著名的例子是蘭花蜘蛛(Thomisus spectabilis),它們能夠改變體色以匹配它們所棲息的花朵,從而更好地捕捉前來訪花的昆蟲。
社會行為的演化也是蜘蛛適應性輻射的一個有趣方面。雖然大多數蜘蛛是獨居的,但一些物種演化出了不同程度的社會性。非洲社會性蜘蛛(Stegodyphus dumicola)就是一個極端的例子,它們形成大型的集體巢穴,共同捕獵和撫育後代。這種社會行為的演化可能是對特定環境壓力的適應,如食物資源的分布或捕食壓力。
蜘蛛在極端環境中的適應也展示了其驚人的適應性輻射能力。例如,喜馬拉雅跳蛛(Euophrys omnisuperstes)被發現生活在海拔6700米的珠穆朗瑪峰上,是已知生活在最高海拔的永久居住動物之一。這種適應展示了蜘蛛如何克服極端的低溫和低氧環境。
蜘蛛的毒素系統也經歷了顯著的適應性輻射。不同物種的蜘蛛演化出了各種複雜的毒素,用於麻痺不同類型的獵物。例如,澳洲的漏斗網蛛(Atrax robustus)演化出了對脊椎動物特別有效的神經毒素,這可能是對其特定生態位中大型獵物的適應。
蜘蛛的適應性輻射還體現在其生殖策略的多樣性上。從複雜的求偶舞蹈到極端的性二型性,再到一些物種中存在的性食同類現象,蜘蛛展示了繁殖策略的廣泛變異。例如,孔雀跳蛛(Maratus volans)雄性的炫耀行為是性選擇壓力下適應性輻射的典型例子。
蜘蛛的適應性輻射過程仍在持續。隨著環境的變化,特別是人類活動對生態系統的影響,蜘蛛正在面臨新的選擇壓力。例如,一些蜘蛛物種已經成功適應了城市環境,如常見的家蛛(Tegenaria domestica)就在人類建築中找到了新的生態位。
研究蜘蛛的適應性輻射不僅有助於我們理解這些生物的演化歷程,還為我們提供了寶貴的視角來理解生物如何應對環境變化和開拓新的生態位。這些知識對於預測和管理生物多樣性,特別是在面對全球環境變化的背景下,具有重要的實際意義。通過深入研究蜘蛛的適應性輻射,我們不僅能更好地理解生物演化的基本原理,還能為保護和管理生態系統提供科學依據。
2.4 全球蜘蛛多樣性
全球蜘蛛多樣性是一個令人驚嘆的生物學現象,反映了這些八腳動物在地球上的廣泛分布和適應能力。目前,科學家已經描述了超過48,000種蜘蛛,但估計實際存在的種類可能超過10萬種。這種豐富的多樣性不僅體現在種類數量上,還表現在形態、行為、生態位和地理分布的巨大變異中。
從地理分布的角度來看,蜘蛛幾乎遍布全球所有的陸地生態系統,從赤道到極地,從海平面到高山,甚至包括一些半水生環境。然而,蜘蛛的多樣性並非均勻分布。熱帶和亞熱帶地區,特別是熱帶雨林,往往擁有最豐富的蜘蛛多樣性。例如,亞馬遜雨林被認為是全球蜘蛛多樣性最高的地區之一,僅在巴西就記錄了超過3,000種蜘蛛。
相比之下,溫帶地區的蜘蛛多樣性相對較低,但仍然相當豐富。例如,整個北美洲記錄了約3,500種蜘蛛,而歐洲則有大約4,000種。即使在氣候條件極為嚴酷的地區,如南極洲,也發現了蜘蛛的蹤跡,儘管種類非常有限。
在形態多樣性方面,蜘蛛展現出驚人的變異。體型大小從體長不到1毫米的微小蜘蛛,如矮蛛科(Micropholcommatidae)的成員,到腿展可達30厘米的巨型鳥蛛,如歌利亞鳥蛛(Theraphosa blondi),範圍極為廣泛。形態變異同樣驚人,從球形的葡萄狀蜘蛛(Patu digua)到棘狀的棘蛛(Gasteracantha),再到扁平如樹皮的樹皮蟹蛛(Bassaniana versicolor),蜘蛛的外形變化無窮。
顏色和圖案的多樣性在蜘蛛中也極為豐富。從鮮豔的孔雀跳蛛(Maratus volans)到幾乎透明的洞穴蜘蛛,蜘蛛的色彩適應反映了它們對不同生態位的適應。一些蜘蛛,如蘭花蟹蛛(Thomisus spectabilis),甚至能夠根據所處環境改變體色,展示了蜘蛛在適應性上的靈活性。
行為多樣性是全球蜘蛛多樣性的另一個重要方面。蜘蛛展現出多種捕獵策略,從織網捕獵到主動狩獵,再到極為特化的捕獵方式。例如,澳大利亞的棒狀蜘蛛(Deinopis spinosa)使用一種獨特的「投網」技術捕獵,而跳蛛科(Salticidae)的成員則依靠敏銳的視力和敏捷的跳躍能力來捕獲獵物。
蜘蛛的築網行為也展現出令人驚嘆的多樣性。從幾何學精確的圓形網,如園蛛科(Araneidae)成員所織造的,到不規則的片狀網,如隱蛛科(Theridiidae)成員所織造的,再到專門用於感知而非捕獵的警戒絲,蜘蛛的網展現出多種形式和功能。一些蜘蛛,如金蛛(Nephila),甚至能夠織造巨大的網,橫跨數米寬的空間。
生態位的多樣性是全球蜘蛛多樣性的另一個重要方面。蜘蛛已經成功適應了各種生態環境,從地下洞穴到樹冠層,從沙漠到熱帶雨林。例如,沙漠中的輪蛛(Cebrennus rechenbergi)能夠通過側翻的方式在沙丘上快速移動,而水蜘蛛(Argyroneta aquatica)則完全適應了水下生活,能夠在水中建造空氣泡作為棲所。
蜘蛛的繁殖策略也展現出豐富的多樣性。從複雜的求偶舞蹈,如跳蛛科成員所展示的,到極端的性食同類現象,如黑寡婦蜘蛛(Latrodectus)中所見的,蜘蛛的繁殖行為變化多端。一些蜘蛛物種展現出高度的親代照顧,如狼蛛科(Lycosidae)的雌性會將卵囊隨身攜帶,而其他物種則幾乎不提供任何親代照顧。
蜘蛛的社會性行為也是其多樣性的一個有趣方面。雖然大多數蜘蛛是獨居的,但一些物種已經演化出不同程度的社會性。例如,非洲社會性蜘蛛(Stegodyphus dumicola)形成大型的集體巢穴,展示了高度的社會合作行為。
全球蜘蛛多樣性的研究不僅有助於我們理解生物多樣性的形成和維持機制,還為生態系統功能和生物地理學研究提供了重要的見解。蜘蛛作為廣泛分布的捕食者,在控制其他節肢動物數量方面扮演著關鍵角色,因此了解蜘蛛的多樣性對於理解和管理生態系統至關重要。
然而,全球蜘蛛多樣性面臨著多重威脅。棲息地破壞、氣候變化、環境污染等因素都可能對蜘蛛多樣性造成負面影響。例如,熱帶雨林的砍伐直接威脅著世界上最豐富的蜘蛛多樣性中心。同時,城市化進程也改變了許多蜘蛛的棲息環境,雖然一些物種成功適應了城市環境,但許多特化的物種可能面臨滅絕風險。
研究和保護全球蜘蛛多樣性不僅對於維護生態系統的平衡至關重要,還可能為人類帶來潛在的利益。例如,蜘蛛毒液中的某些成分被發現具有潛在的藥用價值,而蛛絲的獨特性質也啟發了新材料的研發。因此,保護蜘蛛多樣性不僅是出於生態考慮,也可能為人類的科技和醫學發展帶來意想不到的貢獻。
3. 台灣的蜘蛛概述
台灣的蜘蛛fauna是一個豐富而獨特的生態系統組成部分,反映了該島嶼多樣的地理環境和氣候特徵。作為一個亞熱帶島嶼,台灣擁有從海岸到高山的多樣棲息地,為各種蜘蛛物種提供了理想的生存環境。目前,台灣已記錄的蜘蛛種類超過500種,分屬於40多個科,這個數字隨著研究的深入可能還會增加。
台灣蜘蛛fauna的特點之一是高度的特有性。由於島嶼的地理隔離,許多蜘蛛物種經過長期演化形成了特有種。例如,台灣特有的鐘蛛(Cyclosa taiwanica)就是一個典型案例,這種蜘蛛只在台灣發現。此外,台灣的蜘蛛fauna還包括了許多具有區域代表性的物種,如分布於東亞地區的金蛛(Nephila pilipes),在台灣的低海拔地區常見。
台灣蜘蛛fauna的組成反映了該地區的生物地理特徵,既有來自大陸的種類,也有源自南方熱帶地區的物種。這種多元化的組成使得台灣成為研究蜘蛛生物地理學和演化生物學的重要場所。隨著環境變化和人類活動的影響,台灣的蜘蛛fauna也面臨著保護和管理的挑戰,成為生物多樣性研究和保護的重要對象。
3.1 台灣蜘蛛多樣性
台灣蜘蛛多樣性是該島嶼生態系統中一個引人注目且重要的組成部分。作為一個位於亞熱帶的島嶼,台灣擁有豐富多樣的地理環境和氣候條件,從海岸到高山,從潮濕的雨林到乾旱的河床,為各種蜘蛛種類提供了理想的棲息地。這種環境的多樣性直接反映在台灣蜘蛛fauna的豐富度上。
目前,台灣已記錄的蜘蛛種類超過500種,分屬於40多個科。這個數字雖然相對於全球已知的48,000多種蜘蛛來說似乎不多,但考慮到台灣相對較小的面積,這個多樣性水平實際上是相當可觀的。值得注意的是,隨著研究的深入和新技術的應用,特別是分子生物學方法的使用,這個數字很可能會進一步增加。
台灣蜘蛛多樣性的一個顯著特點是其高度的特有性。由於島嶼的地理隔離,許多蜘蛛種類經過長期演化形成了台灣特有種。例如,台灣特有的鐘蛛(Cyclosa taiwanica)就是一個典型案例。這種蜘蛛只在台灣發現,其獨特的網結構和行為模式反映了長期適應台灣特定環境的結果。類似的例子還有台灣特有的蟹蛛(Tmarus taiwanus)和台灣特有的肖蛸(Psechrus taiwanus)等。
台灣蜘蛛多樣性的另一個特點是其豐富的生態位分布。從海岸到高山,不同的蜘蛛種類適應了不同的生態環境。例如,在台灣的低海拔地區,常見的金蛛(Nephila pilipes)織造巨大的捕獵網,成為當地生態系統中重要的捕食者。而在高山地區,則可以發現一些適應低溫環境的蜘蛛種類,如高山跳蛛(Synagelides taiwanese)。
台灣的蜘蛛多樣性還體現在其豐富的行為和形態變異上。例如,台灣的跳蛛科(Salticidae)成員展示了複雜的求偶行為和色彩變異,如艷彩條紋蛛(Cosmophasis umbratica)雄性的炫耀行為就是一個典型例子。另一方面,一些擬態蜘蛛,如台灣的蟻形蛛(Myrmarachne formicaria),則展示了驚人的形態和行為模仿能力,能夠完美地偽裝成螞蟻。
台灣蜘蛛多樣性的組成反映了該地區的生物地理特徵。台灣的蜘蛛fauna包括了來自大陸的種類,也有源自南方熱帶地區的物種。這種多元化的組成使得台灣成為研究蜘蛛生物地理學和演化生物學的重要場所。例如,台灣的蜘蛛fauna中包含了一些與中國大陸共有的種類,如中國大陸也有分布的異腹蛛(Heteropoda venatoria),同時也包括了一些與菲律賓等東南亞地區共有的種類,如菲律賓也有分布的棕櫚蛛(Heteropoda boiei)。
台灣的蜘蛛多樣性還體現在其豐富的生態功能上。不同的蜘蛛種類在生態系統中扮演著不同的角色。例如,一些大型織網蜘蛛如金蛛(Nephila pilipes)在控制飛蟲數量方面發揮重要作用,而一些地棲蜘蛛如狼蛛科(Lycosidae)的成員則在控制地面昆蟲數量方面起著關鍵作用。這種功能多樣性對於維持生態系統的平衡至關重要。
台灣蜘蛛多樣性的研究還揭示了一些有趣的演化現象。例如,一些研究發現,台灣的某些蜘蛛種類與鄰近地區的近親物種相比,展現出獨特的形態或行為特徵,這可能反映了島嶼環境下的快速演化。這種現象為研究島嶼生物地理學和演化生物學提供了寶貴的素材。
然而,台灣的蜘蛛多樣性也面臨著諸多威脅。棲息地破壞、環境污染、氣候變化等因素都可能對蜘蛛多樣性造成負面影響。例如,台灣的城市化進程改變了許多蜘蛛的棲息環境,雖然一些物種如家蛛(Tegenaria domestica)成功適應了城市環境,但許多特化的物種可能面臨生存壓力。
保護台灣的蜘蛛多樣性不僅對於維護當地生態系統的平衡至關重要,還可能為科學研究和應用帶來潛在的價值。例如,一些台灣特有蜘蛛的毒液成分可能具有潛在的藥用價值,而某些蜘蛛物種的特殊適應能力也可能為生物技術研究提供靈感。
台灣蜘蛛多樣性的研究還涉及了許多有趣的科學問題。例如,台灣的地質歷史如何塑造了當地的蜘蛛fauna?島嶼效應如何影響蜘蛛的演化?這些問題的研究不僅有助於我們理解台灣的生物多樣性,還可能為更廣泛的演化生物學和生態學研究提供重要見解。
此外,台灣蜘蛛多樣性的研究還具有重要的實際應用價值。例如,了解不同蜘蛛種類的生態習性和分布特徑,可以為農業害蟲的生物防治提供重要參考。同時,蜘蛛作為環境變化的敏感指標,對其多樣性的監測也可以為環境保護和生態管理提供重要依據。
3.3A 台灣住家裡的蜘蛛種類?
台灣的住家環境為多種蜘蛛提供了理想的棲息地,這些蜘蛛不僅適應了人類居住空間,還在家庭生態系統中扮演著重要角色。住家蜘蛛的存在雖然有時會引起居民的不安,但實際上它們對控制家居害蟲有著顯著的貢獻。
最常見的住家蜘蛛之一是白額高腳蛛(Heteropoda venatoria),俗稱「大壁虎」。這種體型較大的蜘蛛常見於房屋的牆角、天花板或櫥櫃後方。大魔鬼蛛不織網捕獵,而是靠敏捷的行動能力主動捕捉獵物。它們主要獵食蟑螂、蛾類和其他家居昆蟲,因此在控制室內害蟲方面發揮著重要作用。雖然它們的體型可能令人生畏,但大魔鬼蛛通常會迴避人類,只有在被驚擾時才可能出現防禦行為。
另一種常見的住家蜘蛛是姬蛛(Theridion fatimae),屬於滑腳姬蛛科。這種小型蜘蛛常在房間角落、窗框或傢俱下方織造不規則的網。姬蛛的網雖然看似雜亂,但對捕捉小型飛蟲如蚊子和果蠅非常有效。由於體型小巧且行為隱蔽,姬蛛通常不會引起居民的特別注意,但它們在控制家中小型害蟲方面貢獻顯著。
家幽靈蛛(Pholcus phalangioides),俗稱「長腳蛛」,是另一種在台灣住家中常見的蜘蛛。這種蜘蛛以細長的腿部著稱,常在天花板角落或傢俱下方織造鬆散的網。人面蛛能夠有效捕捉飛蛾、蚊子等飛行昆蟲,同時也會捕食其他小型蜘蛛,在家庭生態系統中扮演著重要的調節者角色。
在台灣的浴室和潮濕區域,常能看到臺灣條紋蚰蜒(Scytodes thoracica)的身影。這種蜘蛛有著獨特的捕獵方式,能夠向獵物噴射黏性的絲液以將其固定。條紋蚰蜒主要捕食小型昆蟲,尤其是在潮濕環境中常見的跳蟲和蠓蚋等。
塔蘭托狼蛛(Lycosa
tarantula)雖然主要棲息在室外,但有時也會進入住家。這種蜘蛛不織網,而是在地面上快速奔跑捕獵。在住家環境中,狼蛛可以有效控制蟑螂、蟋蟀等地面活動的昆蟲。
溫室希蛛(Parasteatoda tepidariorum)是另一種適應良好的住家蜘蛛。這種小型蜘蛛常在窗戶附近或室外植物上織網,能夠捕獲各種小型飛蟲。錢蛛的存在有助於減少蚊蠅等飛行昆蟲進入室內。
在台灣的陽台或庭院中,常能看到嗜水新園蛛(Neoscona
nautica)的大型圓網。雖然圓蛛主要活動在室外,但它們的存在有助於減少飛行昆蟲進入室內。圓蛛的大型蛛網能夠捕獲各種飛蛾、蚊子和其他較大型的飛行昆蟲。
褐條斑蠅虎(Plexippus
paykulli)是一種常見於窗台和牆面的小型蜘蛛。這種蜘蛛不織網,而是依靠敏銳的視力和靈活的跳躍能力捕獵。跳蛛主要捕食蒼蠅、蚊子等小型飛蟲,在室內害蟲控制方面發揮著重要作用。
在一些較老舊或鄉村的住宅中,可能會遇到臺灣蕾蛛(Steatoda grossa)。這種蜘蛛喜歡在黑暗、潮濕的角落築網,其不規則的網能有效捕獲各種爬行昆蟲。蕾蛛在控制蟑螂和螞蟻等家居害蟲方面表現出色。
二角塵蛛(Cyclosa mulmeinensis)雖然主要生活在室外,但有時也會在住家周圍的植物或建築結構上築網。這種蜘蛛的特點是會在網上堆積昆蟲殘骸,形成一條直線,用以掩飾自己的身形。隱圓蛛的存在有助於控制住家周圍的飛行昆蟲數量。
值得一提的是,近年來隨著全球化的發展,一些外來蜘蛛物種也開始在台灣的住家環境中出現。例如,褐隱蛛(Loxosceles reclusa)雖然原產於北美,但已在台灣的一些都市地區被發現。這種蜘蛛喜歡躲藏在衣物堆或紙箱中,其存在提醒我們需要注意外來物種對本地生態系統的潛在影響。
雖然這些蜘蛛在控制家居害蟲方面發揮著重要作用,但它們的存在有時也會引起居民的恐懼或不適。因此,了解這些蜘蛛的生態習性和對環境的積極貢獻,有助於人們以更包容的態度看待這些家居生態系統的重要成員。同時,適當的家居管理,如減少雜物堆積、保持環境整潔等,可以在維持蜘蛛生態效益的同時,減少它們對居住者造成的困擾。
台灣住家蜘蛛的多樣性不僅反映了台灣豐富的生物多樣性,也展示了這些生物對人類居住環境的適應能力。研究和了解這些住家蜘蛛,不僅有助於我們更好地管理家居環境,也為城市生態學和生物多樣性保護提供了重要的研究素材。
3.3B 台灣果園農田裡的蜘蛛種類?
台灣的果園和農田是蜘蛛多樣性的重要棲息地,這些區域為各種蜘蛛提供了豐富的食物來源和多樣的微棲地。蜘蛛在這些農業生態系統中扮演著關鍵的生態角色,尤其是在害蟲控制方面。
在台灣的果園中,最常見的蜘蛛之一是人面蜘蛛(Nephila
pilipes)。這種大型織網蛛常在果樹之間築造巨大的圓網,其金黃色的蛛絲在陽光下格外醒目。金線蛛主要捕食大型飛蟲,如蝴蝶、蛾類和蜜蜂,對控制某些果樹害蟲有顯著作用。例如,在芒果園中,金線蛛能有效捕捉芒果花姬捲葉蛾等害蟲,減少農藥使用需求。
悅目金蛛(Argiope amoena)是另一種在果園中常見的大型織網蛛。這種蜘蛛以其鮮豔的黑黃相間體色和特殊的「X」形穩定帶蛛網而聞名。銀腹蛛主要分布在果園的邊緣地帶,能夠捕獲多種飛行昆蟲,包括果實蠅和小型蛾類,對果樹病蟲害防治有重要貢獻。
在低矮的果樹或灌木叢中,常能發現塔蘭托狼蛛(Lycosa tarantula)的蹤跡。這種地棲性蜘蛛不織網,而是在地面上快速奔跑捕獵。狼蛛主要捕食各種地面活動的昆蟲,如蝗蟲、蟋蟀和某些甲蟲幼蟲,對控制土壤害蟲有很大幫助。在柑橘園中,狼蛛能有效捕食影響根系的害蟲。
姬蛛(Theridion fatimae)是果園中常見的小型蜘蛛。這種蜘蛛常在果樹枝葉間織造不規則的網,主要捕食小型飛蟲如蚜蟲和葉蟬。在葡萄園中,姬蛛對控制葡萄蚜蟲等微小害蟲有重要作用。
在果園的地面層,古氏棘蛛(Gasteracantha kuhlii)常在低矮植物間築網。這種蜘蛛以其獨特的盾形外觀而聞名,能夠有效捕獲各種低飛昆蟲。在草莓園中,盾蛛對控制草莓小葉蜂等低飛害蟲有一定效果。
轉向農田環境,我們可以發現不同的蜘蛛群落組成。在水稻田中,日本長腳蛛(Tetragnatha maxillosa)是最常見的蜘蛛之一。這種長腳蛛常在稻株間織造水平網,主要捕食飛蝨、葉蟬等水稻害蟲。稻蛛的存在大大減少了農民對化學農藥的依賴。
擬水狼蛛(Pirata subpiraticus)是另一種常見於水田的蜘蛛。這種半水棲蜘蛛能在水面上行走,主要捕食水生昆蟲和落入水中的陸生昆蟲。在水稻生長的不同階段,臺灛蛛都能發揮重要的害蟲控制作用。
在蔬菜園中,嗜水新園蛛(Neoscona nautica)是常見的大型織網蛛。圓蛛每晚都會重新織造完整的圓網,能夠有效捕獲各種飛行昆蟲。在茄子園和青椒園中,圓蛛對控制夜蛾類害蟲特別有效。
斜紋貓蛛(Oxyopes sertatus)是蔬菜園中常見的遊獵性蜘蛛。這種蜘蛛不織網,而是在植物上快速移動捕捉獵物。蠅虎對控制蚜蟲、葉蟬等小型害蟲有顯著效果,在葉菜類種植中尤為重要。
在一些特殊的農業環境中,如設施農業,我們可以發現適應這種環境的特殊蜘蛛種類。例如,在溫室中常見褐條斑蠅虎(Plexippus
paykulli)。這種視力敏銳的小型蜘蛛能在有限的空間內有效捕獲各種小型害蟲,對於溫室害蟲的綜合防治有重要意義。
在果園和農田的邊緣地帶,細紋貓蛛(Oxyopes macilentus)是常見的蜘蛛種類。這種蜘蛛具有良好的偽裝能力,能在植物上靜待獵物接近。肖蛸對控制果園和農田周邊的潛在害蟲入侵有重要作用。
值得注意的是,隨著有機農業的發展,一些對環境敏感的蜘蛛種類開始在台灣的農業生態系統中重新出現。例如,三角蟹蛛(Thomisus labefactus)在有機果園中的數量明顯增加。這種蛛不織網,而是在花朵上埋伏獵物,對控制授粉昆蟲數量有重要作用。
在一些特殊作物的種植區,如茶園,我們可以發現特化的蜘蛛群落。例如,白斑獵蛛(Evarcha albaria)是茶園中常見的跳蛛,它們主要捕食茶小綠葉蟬等茶樹特有害蟲。
農田和果園的蜘蛛不僅在害蟲控制方面發揮作用,還是評估農業生態系統健康的重要指標。例如,斜紋貓蛛(Oxyopes sertatus)對農藥敏感,其數量可以反映農田生態環境的質量。
這些蜘蛛種類的存在和數量變化,不僅反映了台灣農業生態系統的多樣性和複雜性,也為農業害蟲的生物防治提供了重要支持。了解和保護這些蜘蛛,對於發展可持續農業實踐、減少化學農藥使用、維護農業生態平衡具有重要意義。同時,這些蜘蛛的研究也為台灣的生物多樣性保護和農業生態學研究提供了豐富的素材。
3.3C 台灣丘陵森林裡的蜘蛛種類?
台灣的丘陵森林是蜘蛛多樣性的重要棲息地,這裡複雜的生態環境為各種蜘蛛提供了豐富的微棲地和食物資源。從地表到樹冠,不同的森林層次都有其特有的蜘蛛群落,展現了令人驚嘆的生態適應性和多樣性。
在森林地表層,塔蘭托狼蛛(Lycosa
tarantula)是最常見的蜘蛛之一。這種大型地棲蜘蛛不織網,而是利用其敏捷的身手在落葉層中奔跑捕獵。狼蛛主要捕食各種地面昆蟲,如甲蟲、蟋蟀和蚱蜢,在維持森林地表生態平衡中扮演重要角色。在台灣中部的八仙山自然保護區,研究人員發現狼蛛的數量與地表昆蟲的多樣性呈正相關,反映了其在生態系統中的關鍵地位。
稍高於地表的草本層,三角蟹蛛(Thomisus labefactus)是一種常見的伏擊型捕食者。這種蜘蛛通常棲息在花朵或葉片上,憑藉其出色的偽裝能力等待獵物靠近。蟹蛛在控制授粉昆蟲數量方面發揮著微妙的平衡作用,間接影響了森林植物的繁殖成功率。在台灣東部的花蓮林區,研究發現蟹蛛的存在顯著影響了某些蝴蝶蘭的授粉效率。
在灌木層,姬蛛(Theridion fatimae)是常見的小型織網蛛。這種蜘蛛在枝葉間織造不規則的網,主要捕食小型飛蟲如蚊子和果蠅。姬蛛的存在有助於控制可能傳播植物病毒的小型昆蟲,對維護森林健康具有重要意義。在台中大坑風景區的次生林中,研究人員觀察到姬蛛數量與小型昆蟲的種群密度呈顯著負相關。
樹幹是另一個重要的蜘蛛棲息微環境。臺灣樹蛛(Heteropoda venatoria)常見於較粗大的樹幹上,這種大型蜘蛛不織網,而是利用其敏捷的身手在樹幹表面捕獵各種昆蟲。樹蛛在控制樹皮甲蟲等森林害蟲方面發揮重要作用。在台南龍崎牛埔自然保留區,研究發現樹蛛的存在顯著降低了某些樹種的樹皮受損程度。
在較高的樹冠層,人面蜘蛛(Nephila pilipes)是最引人注目的蜘蛛之一。這種大型織網蛛在樹木之間織造巨大的圓網,主要捕食大型飛蟲如蛾類和蝴蝶。金線蛛的存在不僅控制了某些森林害蟲的數量,其龐大的蛛網還為其他小型生物如某些鳥類提供了築巢材料。在台北陽明山國家公園,研究人員發現金線蛛的分佈與某些鳥類的築巢地點呈現顯著相關性。
在森林的開闊地帶或邊緣,悅目金蛛(Argiope
amoena)常常可見。這種蜘蛛以其鮮豔的體色和特殊的「X」形穩定帶蛛網而聞名。銀腹蛛主要捕食各種中小型飛蟲,在控制森林邊緣害蟲入侵方面發揮著重要作用。在宜蘭福山植物園,研究發現銀腹蛛的存在顯著降低了外來害蟲對原生植物的危害。
在森林的陰暗潮濕處,如岩石縫隙或倒木下,常能發現臺灣皿蛛(Atypus karschi)的蹤跡。這種原始蜘蛛構築獨特的襪狀巢穴,是台灣少數具有長壽命的蜘蛛物種之一。皿蛛的存在被認為是森林生態系統穩定性的一個重要指標。在台東知本森林遊樂區,長期監測顯示皿蛛數量的穩定與森林生態系統的整體健康狀況高度相關。
森林中的特殊微棲地,如樹洞或枯枝,也孕育了獨特的蜘蛛種類。例如,臺灣捲葉蛛(Phonognatha fumosa)常在這些環境中築巢。這種蜘蛛利用捲曲的樹葉作為隱蔽所,展現了高度的環境適應能力。在台中大雪山森林遊樂區,研究人員發現捲葉蛛的分佈與森林的樹種多樣性呈現正相關。
在森林的中下層,古氏棘蛛(Gasteracantha kuhlii)以其獨特的外形吸引了許多研究者的注意。這種蜘蛛的腹部呈現盾牌狀,常在樹枝間織造小型但堅韌的圓網。盾蛛在捕捉小型飛蟲方面表現出色,對維持森林昆蟲群落的平衡具有重要作用。在新竹觀霧森林遊樂區,生態學家發現盾蛛的數量與某些植物的結實率存在正相關,暗示了其在森林生態系統中的複雜作用。
台灣丘陵森林中還有一些特化的蜘蛛種類,如臺灣葉蛛(Poltys illepidus)。這種蜘蛛白天時會將身體捲縮,完美地模仿枯枝或樹芽,展現了驚人的擬態能力。夜間,葉蛛會展開身體,織網捕獵。在台南曾文水庫集水區的森林中,研究人員發現葉蛛的存在與某些夜行性昆蟲的數量控制有密切關係。
值得注意的是,台灣丘陵森林中的蜘蛛群落還包括一些稀有或特有種。例如,臺灣細蛛(Ariamnes cylindrogaster)是一種形態奇特的蜘蛛,其細長的身體能夠完美地隱藏在樹枝或藤蔓之間。這種蜘蛛的發現為台灣的生物多樣性研究提供了重要資料。在屏東茂林國家風景區的森林中,細蛛的存在被視為生態系統健康的重要指標之一。
這些豐富多樣的蜘蛛種類不僅展示了台灣丘陵森林生態系統的複雜性,也反映了蜘蛛在維持生態平衡中的重要作用。從控制昆蟲數量到影響植物授粉和繁殖,再到作為生態系統健康的指標,森林蜘蛛在多個層面上參與並影響著生態過程。深入研究這些蜘蛛,不僅能增進我們對台灣生物多樣性的理解,還能為森林生態系統的保護和管理提供重要的科學依據。
3.3C 台灣高山森林裡的蜘蛛種類?
台灣的高山森林是一個獨特而多樣的生態系統,為許多特化的蜘蛛種類提供了棲息地。這些高海拔環境因其特殊的氣候條件和植被結構,孕育了一系列適應高山生活的蜘蛛群落。從亞高山針葉林到高山草原,不同的生態區域都有其特有的蜘蛛種類。
在台灣高山森林的地表層,臺灣高山狼蛛(Pardosa pseudoannulata)是一種常見的地棲蜘蛛。這種蜘蛛適應了高海拔的寒冷環境,能在低溫條件下保持活躍。它們主要在落葉層和岩石縫隙中活動,捕食各種小型節肢動物。在玉山國家公園的高山森林區,研究人員發現高山狼蛛的數量與地表小型無脊椎動物的多樣性呈正相關,反映了其在高山生態系統食物網中的重要地位。
高山森林的灌木層中,臺灣高山蟹蛛(Xysticus ephippiatus)是一種特化的伏擊型捕食者。這種蜘蛛能夠在寒冷的環境中保持活力,常棲息在高山植物的花朵或葉片上。蟹蛛在控制高山植物授粉昆蟲數量方面發揮著微妙的平衡作用。在雪山地區的高山草甸中,研究發現高山蟹蛛的存在影響了某些高山植物的授粉效率,進而影響了植物群落的組成。
在高山森林的針葉樹上,臺灣高山圓蛛(Araneus
ventricosus)是一種適應高海拔環境的織網蛛。這種蜘蛛能夠在寒冷和強風的條件下織造堅韌的圓網,主要捕食高空飛行的昆蟲。在太魯閣國家公園的高山區,研究人員觀察到高山圓蛛的網絲結構比低海拔地區的同類蜘蛛更為堅固,這被認為是對高山強風環境的適應。
高山森林中的特殊微棲地,如岩石縫隙或苔蘚層,孕育了一些獨特的蜘蛛種類。例如,臺灣高山皿蛛(Atypus karschi)常在這些環境中築巢。這種原始蜘蛛在高山環境中展現了驚人的適應能力,能夠在寒冷乾燥的條件下生存。在合歡山區,長期監測顯示高山皿蛛的分佈與高山生態系統的穩定性高度相關。
在高山森林的樹幹上,臺灣高山樹蛛(Heteropoda venatoria)展現了對寒冷環境的適應。這種大型蜘蛛不織網,而是利用其敏捷的身手在樹幹表面捕獵各種昆蟲。在大霸尖山地區,研究發現樹蛛在控制某些高山樹木害蟲方面發揮著重要作用,對維護高山森林生態平衡具有重要意義。
高山草原是另一個重要的蜘蛛棲息環境。在這裡,臺灣高山跳蛛(Evarcha albaria)是一種常見的捕食者。這種小型蜘蛛具有出色的跳躍能力和敏銳的視力,能夠在開闊的草地上高效捕獵。在奇萊山區的高山草原中,研究人員發現高山跳蛛的數量與某些高山昆蟲種群的波動密切相關。
在高山森林的樹冠層,臺灣高山姬蛛(Theridion pinastri)展現了對高海拔環境的獨特適應。這種小型蜘蛛在針葉樹的枝條間織造不規則的網,主要捕食小型飛蟲。在台灣中央山脈的高山地區,研究發現高山姬蛛的存在對控制某些針葉樹害蟲具有重要作用。
高山森林中的一些特殊環境,如高山湖泊周圍,也孕育了獨特的蜘蛛種類。例如,臺灣高山漁蛛(Dolomedes sulfureus)常見於高山湖泊和溪流周圍。這種半水棲蜘蛛能夠在水面上行走,捕食水生昆蟫和掉落水面的陸生昆蟲。在七彩湖周圍,研究人員發現漁蛛在維持高山水域生態平衡方面扮演著重要角色。
值得注意的是,台灣高山森林中還存在一些稀有或特有的蜘蛛種類。例如,臺灣高山細蛛(Ariamnes cylindrogaster)是一種形態特殊的蜘蛛,其細長的身體能夠完美地隱藏在高山植物的枝條間。在玉山園區的高山森林中,細蛛的發現為台灣高山生態系統的生物多樣性研究提供了重要資料。
高山森林的極端環境也促使一些蜘蛛發展出特殊的生存策略。例如,某些高山蜘蛛物種能夠產生類似於防凍蛋白的物質,使其能在接近冰點的溫度下生存。在雪山東峰地區,研究人員發現某些高山蜘蛛即使在冬季也能保持活動,這種適應能力在全球蜘蛛種類中極為罕見。
台灣高山森林中的蜘蛛不僅展現了驚人的適應能力,也在生態系統中扮演著關鍵角色。從控制昆蟲數量到影響植物授粉,再到作為環境變化的指示物種,這些高山蜘蛛以多種方式參與並影響著高山生態系統的運作。
例如,在玉山國家公園的研究中,科學家發現某些高山蜘蛛物種的分佈範圍正在隨著氣候變暖而逐漸上移。這一現象不僅反映了氣候變化對高山生態系統的影響,也為研究全球變暖對生物多樣性的影響提供了重要案例。
此外,高山蜘蛛的研究也為生物地理學提供了寶貴資料。台灣高山森林中的一些特有蜘蛛種類,可能是冰河時期遺留下來的孑遺物種。通過研究這些蜘蛛的遺傳特徵和分佈模式,科學家們能夠重建台灣島的生物地理歷史,進而深化我們對東亞地區生物多樣性形成過程的理解。
高山森林蜘蛛的研究不僅具有重要的科學價值,也為高山生態系統的保護和管理提供了重要依據。這些蜘蛛的存在和數量變化,可以作為評估高山生態系統健康狀況的重要指標。因此,持續監測和研究這些高山蜘蛛,對於制定有效的高山生態系統保護策略具有重要意義。
3.3E 台灣住家周邊與公園最常見的蜘蛛種類15種?
台灣住家周邊與公園環境中常見的蜘蛛種類豐富多樣,這些蜘蛛不僅適應了人類活動頻繁的區域,還在城市生態系統中扮演著重要角色。以下介紹15種最常見的蜘蛛,包括它們的外觀特徵、棲息地選擇、繁殖與生活習性等方面:
1.
白額高腳蛛(Heteropoda venatoria) 外觀特徵:體長可達3-4公分,體色為黃褐色或灰褐色,腹部有不規則的深色斑紋。 棲息地選擇:常見於住家內部,如牆角、天花板或櫥櫃後方。 繁殖與生活習性:雌蛛會攜帶圓形的卵囊,孵化期約一個月。夜行性,主動獵捕蟑螂等昆蟲。
2.
家幽靈蛛(Pholcus phalangioides) 外觀特徵:體型細長,腿部特別長,體長約7-8毫米。 棲息地選擇:喜歡在室內陰暗角落築網,如浴室、地下室等。 繁殖與生活習性:雌蛛將卵囊含在口器附近。當受到驚擾時,會快速震動身體。
3.
姬蛛(Theridion fatimae) 外觀特徵:體型小,約3-5毫米,腹部呈圓形,常有明顯的花紋。 棲息地選擇:常在窗框、傢俱縫隙等處築不規則網。 繁殖與生活習性:雌蛛會在網中央照顧卵囊和幼蛛。主要捕食小型飛蟲。
4.
褐條斑蠅虎(Plexippus
paykulli) 外觀特徵:體長約8-10毫米,有明顯的大眼睛,體色多樣。 棲息地選擇:常見於陽台、庭院的植物上或牆面。 繁殖與生活習性:不織網,靠跳躍捕獵。雄蛛會進行複雜的求偶舞蹈。
5.
嗜水新園蛛(Neoscona
nautica) 外觀特徵:體長約15-20毫米,腹部呈卵圓形,常有明顯的十字花紋。 棲息地選擇:常在庭院或公園的樹木間織大型圓網。 繁殖與生活習性:每晚重建蛛網,白天躲藏在葉片捲曲處。
6.
悅目金蛛(Argiope
amoena) 外觀特徵:體長可達25-30毫米,腹部有明顯的黑黃相間條紋。 棲息地選擇:喜歡在開闊地帶如庭院、公園草地間築網。 繁殖與生活習性:蛛網中常有醒目的「X」形穩定帶。卵囊呈梨形,懸掛在植物上。
7.
塔蘭托狼蛛(Lycosa
tarantula) 外觀特徵:體型強健,體長約20-25毫米,體色多為褐色。 棲息地選擇:常見於地面、落葉層或草叢中。 繁殖與生活習性:雌蛛會將卵囊附在紡織器上攜帶。夜行性,主動獵捕地面昆蟲。
8.
斜紋貓蛛(Oxyopes sertatus) 外觀特徵:體長約10-12毫米,腹部細長,常有縱向條紋。 棲息地選擇:多見於灌木或草本植物上。 繁殖與生活習性:不織網,在植物間快速移動捕食。雌蛛將卵囊藏在葉片捲起的空間內。
9.
古氏棘蛛(Gasteracantha kuhlii) 外觀特徵:腹部呈盾形,有明顯的刺狀突起,體色鮮艷。 棲息地選擇:常在低矮灌木間築小型圓網。 繁殖與生活習性:網上常有醒目的絲帶裝飾。卵囊呈圓盤狀,藏在樹皮縫隙中。
10.
三角蟹蛛(Thomisus labefactus) 外觀特徵:體型扁平,前兩對腳特別長,體色可隨環境變化。 棲息地選擇:常見於花朵或葉片上。 繁殖與生活習性:不織網,採取伏擊式捕獵。雌蛛會在捲起的葉片中產卵。
11.
臺灣姥蛛(Nephila
pilipes) 外觀特徵:雌蛛體長可達5公分,金黃色體表有黑色斑點。 棲息地選擇:在開闊處如庭院、公園邊緣織造大型圓網。 繁殖與生活習性:雌雄體型差異極大。雄蛛常停留在雌蛛網緣。
12.
臺灣條紋蚰蜒(Scytodes thoracica) 外觀特徵:體長約5-7毫米,體表有複雜的條紋圖案。 棲息地選擇:常見於室內陰暗潮濕處。 繁殖與生活習性:能夠噴射黏性絲液捕獵。雌蛛將卵囊含在口器下。
13.
溫室希蛛(Parasteatoda tepidariorum) 外觀特徵:體長約5-6毫米,腹部圓形,常有不規則斑點。 棲息地選擇:常在室外角落或植物間築不規則網。 繁殖與生活習性:雌蛛在網中央照顧多個梨形卵囊。
14.
臺灣蛇蛛(Tetragnatha maxillosa) 外觀特徵:體型細長,螯肢特別發達,體長約10-15毫米。 棲息地選擇:常見於水邊植物或建築物外牆。 繁殖與生活習性:織造水平或斜向的圓網。白天常將身體貼附在枝條上休息。
15.
臺灣寡婦蛛(Latrodectus
elegans) 外觀特徵:雌蛛體長約10毫米,腹部黑色有紅色斑點。 棲息地選擇:喜歡在陰暗處如石縫、木堆下築網。 繁殖與生活習性:織造不規則網。雌蛛會守護球形的白色卵囊。咬傷可能造成嚴重症狀,需小心避免接觸。
1.
大魔鬼蛛 (Heteropoda venatoria)
2.
人面蛛 (Pholcus phalangioides)
3.
姬蛛 (Theridion sp.)
4.
跳蛛
(Plexippus paykulli)
5.
圓蛛
(Neoscona nautica)
6.
銀腹蛛
(Argiope amoena)
7.
狼蛛
(Lycosa sp.)
8.
蠅虎 (Oxyopes sertatus)
9.
盾蛛 (Gasteracantha kuhlii)
10.
蟹蛛 (Thomisus labefactus)
11.
金線蛛
(Nephila pilipes)
12.
條紋蚰蜒 (Scytodes thoracica)
13.
錢蛛 (Parasteatoda tepidariorum)
14.
蛇蛛 (Tetragnatha maxillosa)
15.
黑寡婦蛛
(Latrodectus elegans)
3.q 台灣都市環境中的蜘蛛
台灣的都市環境為蜘蛛提供了一個獨特而複雜的生態系統。隨著城市化的快速發展,許多蜘蛛種類已經適應並繁衍在這種人為主導的環境中。台灣的都市蜘蛛不僅展現了驚人的適應能力,還在城市生態系統中扮演著重要角色。
在台北、高雄等大都市中,常見的都市蜘蛛包括 白額高腳蛛(Heteropoda venatoria)、溫室希蛛(Parasteatoda tepidariorum)和黑斑園蛛(Araneus mitificus)等。這些蜘蛛已經成功地將人類建築和都市基礎設施轉化為自己的棲息地。例如,白額高腳蛛經常出現在建築物的角落、天花板或牆壁上。它們利用這些位置作為隱蔽處,同時捕食室內外的小型昆蟲。這種適應不僅展現了蜘蛛的生存智慧,也為都市環境中的害蟲控制做出了貢獻。
都市公園和綠地是台灣城市蜘蛛的重要棲息地。在台北的大安森林公園或高雄的中央公園等地,可以觀察到多種蜘蛛活動。例如,人面蜘蛛(Nephila pilipes)常在樹木間織造大型圓網,這些網不僅是捕食昆蟲的工具,還為公園增添了自然美感。同時,這些綠地為蜘蛛提供了都市中難得的自然棲息環境,維持了城市生物多樣性。
台灣都市中的人工建築為某些蜘蛛種類創造了新的生態位。例如,台灣姬蛛常在窗框、屋簷下或陽台欄杆等處築網。這些位置不僅提供了穩定的結構支撐,還能利用人工照明吸引來的昆蟲作為食物來源。這種適應性使得台灣姬蛛成為都市環境中最常見的蜘蛛之一。
都市化帶來的微氣候變化也影響了蜘蛛的分布和行為。城市熱島效應使得都市環境的溫度通常高於周邊鄉村地區。這種溫度差異可能導致某些耐熱的蜘蛛種類在都市中更為常見。例如,在台中市區,常見的人面蜘蛛似乎比郊區更能適應高溫環境,它們能在陽光直射的建築外牆上織網捕食。
台灣都市環境中的蜘蛛還面臨著獨特的挑戰和機遇。例如,頻繁的人為干擾可能會破壞蜘蛛的網。然而,有些蜘蛛種類已經適應了這種情況,能夠快速重建或修復被破壞的網。此外,都市環境中的光污染雖然可能干擾某些蜘蛛的自然行為,但也為夜行性蜘蛛提供了額外的捕食機會,因為許多昆蟲會被人工照明吸引。
都市蜘蛛在台灣城市生態系統中扮演著重要角色。作為捕食者,它們有助於控制蚊子、蒼蠅等城市害蟲的數量。例如,在台南市的一項研究中發現,屋簷下的姬蛛網能有效捕捉登革熱病媒蚊。這種自然的害蟲控制不僅對維護城市環境衛生有所幫助,還可能減少殺蟲劑的使用,從而降低環境污染。
然而,台灣都市環境中的蜘蛛也面臨著一些威脅。過度使用殺蟲劑、頻繁的清潔和維護活動,以及棲息地的碎片化都可能對蜘蛛種群產生負面影響。例如,在台北市的一些新開發區域,由於綠地減少和殺蟲劑使用增加,某些原本常見的蜘蛛種類變得罕見。
台灣的都市規劃者和生態學家正在努力創造更加友善蜘蛛的城市環境。例如,在新北市的某些社區,通過減少殺蟲劑使用和增加綠化面積,成功地提高了當地蜘蛛的多樣性。這不僅有利於生態平衡,還為市民提供了觀察和了解這些有趣生物的機會。
都市蜘蛛還為台灣的環境教育提供了寶貴的資源。許多學校和社區組織利用都市中常見的蜘蛛進行自然教育活動。例如,在台中市的一些小學,教師會帶領學生觀察校園內的蜘蛛,教導他們識別不同種類,並了解蜘蛛在生態系統中的作用。這種親近自然的教育方式不僅增加了學生的生態知識,還有助於消除對蜘蛛的恐懼和誤解。
台灣都市環境中的蜘蛛多樣性也反映了城市的生態健康狀況。一些研究表明,蜘蛛的種類和數量可以作為評估都市生態系統質量的指標。例如,在高雄市的一項調查中發現,擁有更多綠地和水體的區域通常有更豐富的蜘蛛多樣性。這為城市規劃者提供了有價值的參考,幫助他們在都市發展中更好地兼顧生態保育。
隨著城市化的持續推進,台灣的都市蜘蛛正在不斷適應和進化。一些原本生活在自然環境中的蜘蛛種類正逐漸適應都市生活。例如,原本主要分布在低海拔森林的台灣爬牆蛛(Uloborus formosanus),近年來已經在台北市的一些老舊建築外牆上被發現。這種現象不僅展示了蜘蛛的適應能力,也為研究城市化對生物進化的影響提供了有趣的案例。
都市蜘蛛在台灣城市景觀中也扮演著獨特的美學角色。例如,在秋季的清晨,人們常能在公園或庭院中看到被露水裝飾的蜘蛛網,這種自然景觀為繁忙的都市生活增添了一絲詩意。一些藝術家和攝影愛好者甚至專門以都市蜘蛛為主題創作,展現城市與自然和諧共存的美感。
3.ab 台灣丘陵邊緣常見的蜘蛛?
台灣低海拔丘陵樹林中的蜘蛛群落在面對人為干擾時展現出了獨特的適應性和豐富的多樣性。這種環境通常位於都市邊緣或鄉村地區,是自然生態系統和人類活動交匯的地帶,為研究蜘蛛對人為影響的適應提供了理想的場所。
在這樣的環境中,我們可以觀察到多種蜘蛛類群的共存。其中,園蛛科(Araneidae)的成員往往最為顯眼。台灣絲絨金蛛(Nephila pilipes)是一個典型的代表,這種大型蜘蛛能在樹木之間織造巨大的圓形網。儘管人為活動可能會破壞它們的網,但這些蜘蛛表現出了驚人的恢復能力,能夠迅速重建或修復被破壞的網。此外,溫室希蛛(Parasteatoda tepidariorum)也常見於這種環境,它們傾向於在建築物的角落或植物枝葉間織網,展現出對人為結構的良好適應。
狼蛛科(Lycosidae)的成員在這種環境中也相當常見。它們不織網,而是在地面上活動獵食。台灣姥姥蛛(Lycosa formosana)經常可以在落葉層或低矮植被中發現。這些蜘蛛似乎能夠適應人為干擾帶來的環境變化,在修剪過的草地和自然生長的灌木叢中都能生存。
跳蛛科(Salticidae)的多樣性在這種環境中尤為豐富。這些色彩斑斕、行為有趣的小型蜘蛛能夠在各種微棲息地中生存。例如,台灣雙斑跳蛛(Plexippus
setipes)常見於建築物外牆或樹幹上,它們敏捷的跳躍和獨特的捕獵方式使其成為觀察的理想對象。人為環境中的多樣表面結構似乎為跳蛛提供了豐富的獵場。
人為干擾對蜘蛛群落的影響是多方面的。一方面,某些干擾活動如除草、修剪可能會破壞蜘蛛的棲息地。但另一方面,這些活動也可能創造新的生態位。例如,經常修剪的草地可能不適合某些織網蜘蛛,但卻為地棲性的狼蛛提供了理想的獵場。此外,人工照明雖然可能干擾某些夜行性蜘蛛的自然行為,但也為其他種類提供了額外的捕食機會,因為許多昆蟲會被燈光吸引。
在這種環境中,我們還可以觀察到蜘蛛對人造結構的適應。例如,台灣鳳蝶蛛(Argiope aemula)經常在花園植物或建築物周圍的開放空間織網。它們似乎能夠利用人造結構作為網的支撐點,有時甚至會在晾衣架或陽台欄杆上築網。這種適應性不僅展示了蜘蛛的靈活性,也為居民提供了近距離觀察這些有趣生物的機會。
人為干擾還可能影響蜘蛛的食物來源。例如,果園或菜園中使用的農藥可能減少昆蟲數量,從而影響依賴這些昆蟲為食的蜘蛛。然而,某些蜘蛛物種表現出了適應這種變化的能力。台灣單線長約蛛(Tetragnatha maxillosa)就常見於農業環境中,它們能夠在作物行間織網,捕食未被農藥消滅的小型昆蟲。
值得注意的是,這種環境中的蜘蛛群落也受到季節變化的影響。在台灣,春季和秋季通常是蜘蛛活動的高峰期。這與人類活動的季節性模式相互作用,例如,春季園藝活動的增加可能為某些蜘蛛創造新的棲息地,而秋季落葉的堆積則為地棲蜘蛛提供了良好的隱蔽所。
此外,這種環境中的蜘蛛還面臨來自入侵種的競爭。例如,入侵的黃褐歡蛛(Latrodectus geometricus)在台灣的低海拔地區逐漸變得常見。這種蜘蛛適應性強,能夠在人為環境中迅速建立種群,可能對本地蜘蛛種類造成競爭壓力。
人為干擾也可能改變蜘蛛的行為模式。例如,在頻繁受到干擾的區域,一些蜘蛛物種可能會變得更加警惕或更頻繁地重建網。台灣光喙蛛(Araneus mitificus)就展現出了這種適應性,它們通常會在傍晚織網,並在清晨拆除,以避免白天的人為干擾。
這種環境中的蜘蛛群落對生態系統服務具有重要意義。它們不僅控制昆蟲數量,維持生態平衡,還可能幫助減少某些有害昆蟲的繁殖。例如,在台灣南部的一項研究中發現,住家周圍的蜘蛛群落對控制登革熱病媒蚊的數量有一定作用。
然而,人們對蜘蛛的態度也會影響這些生物在人為環境中的生存。許多人對蜘蛛存在恐懼或反感,可能會主動清除家中或院子裡的蜘蛛。這種行為可能導致局部區域的蜘蛛多樣性下降。因此,提高公眾對蜘蛛生態價值的認識變得尤為重要。
在台灣低海拔丘陵樹林有住家人為干擾的環境中,蜘蛛群落展現了複雜的生態動態。它們不僅適應了人為環境的挑戰,還在某些方面受益於這種環境。這種人與自然的互動為研究城市化對生物多樣性的影響提供了寶貴的見解,同時也突顯了在城市規劃和環境管理中考慮生物多樣性的重要性。
台灣溪流上游的濱溪環境是一個獨特的生態系統,為多種蜘蛛提供了理想的棲息地。這種環境通常包括溪流、岩石、濱溪植被和少量的人為干擾(如農家),形成了豐富多樣的微棲息地。在這樣的環境中,常見的蜘蛛包括以下幾種:
縱條銀鱗蛛(Leucauge magnifica)是在這種環境中最常見的蜘蛛之一。這種蜘蛛以其美麗的銀綠色體色和精巧的圓形網而聞名。它們常在溪流上方的植被間築網,利用水面上飛行的昆蟲作為食物來源。單線階蛛對濕度要求較高,因此在濱溪環境中特別常見。
人面蜘蛛(Nephila pilipes)也是濱溪環境中的常見物種。這種大型蜘蛛喜歡在樹木或灌木之間築造大型的黃金色蛛網。它們通常選擇較為開闊的空間,如溪流兩岸的空地或農家附近的樹木間。金線蛛的存在往往標誌著良好的生態環境。
臺灣銀腹蛛(Argiope amoena)是另一種在濱溪環境中常見的蜘蛛。這種蜘蛛以其明顯的黑黃相間的體色和特徵性的「X」形網飾而易於識別。它們常在草叢或低矮灌木間築網,捕食各種飛行昆蟲。
溪流上游環境中還經常可以看到狼蛛科的物種,如臺灣狼蛛(Lycosa coelestis)。這些蜘蛛不織網,而是在地面上活動獵食。它們常在溪邊的石頭下或落葉堆中棲息,捕食各種小型節肢動物。
另一種常見的是姬蛛(Oxyopes sertatus)。這種蜘蛛體型較小,常在低矮的植被上活動。它們是敏捷的獵手,能夠快速奔跑和跳躍來捕捉獵物。
在較為潮濕的區域,如靠近溪流的岩石或樹幹上,可能會發現臺灣壁蛛(Heteropoda venatoria)。這種大型的遊蛛不織網,而是主動獵捕獵物。它們的扁平體型使其能夠輕易躲藏在狹窄的縫隙中。
此外,在濱溪植被中,常常可以看到園蛛科的物種,如臺灣圓蛛(Neoscona punctigera)。這些蜘蛛織造典型的輪輻狀蛛網,常見於草叢或低矮灌木間。
值得注意的是,具體的蜘蛛種類可能會因季節、海拔高度和微棲息地的不同而有所變化。此外,一些稀有或特有種可能也會在這種環境中出現,但需要更專業的調查才能發現。這些蜘蛛共同構成了台灣溪流上游濱溪環境的豐富生態系統,在維持生態平衡和控制昆蟲數量方面發揮著重要作用。
3.2 台灣主要的.常見的的蜘蛛科?特徵?棲地?習性?
台灣是一個生物多樣性豐富的島嶼,其獨特的地理位置和多樣化的生態環境為許多蜘蛛物種提供了理想的棲息地。在台灣的蜘蛛fauna中,有幾個主要的科別特別常見且具有代表性。
首先,我們來談談園蛛科(Araneidae)。這個科的蜘蛛在台灣非常普遍,其中最為人熟知的莫過於金線蛛屬(Nephila)的人面蜘蛛(Nephila
pilipes)。這種蜘蛛體型巨大,雌蛛體長可達5公分,而雄蛛則相對較小。金線蛛以其金黃色的絲線而聞名,它們通常在開闊的空間如森林邊緣或庭園中織造巨大的圓形網。這種蜘蛛具有顯著的性二型性,雌蛛不僅體型更大,而且色彩也更為艷麗。金線蛛主要以飛蛾和其他飛行昆蟲為食,它們的網具有極強的黏性和韌性,能夠捕捉到較大型的獵物。
另一個在台灣常見的蜘蛛科是狼蛛科(Lycosidae)。這類蜘蛛不織網捕獵,而是主動在地面上追捕獵物。它們通常體型中等,具有發達的視覺系統,能夠在夜間高效地捕獵。狼蛛科的一個顯著特徵是雌蛛會將卵囊附著在腹部下方攜帶,孵化後的幼蛛會爬到母蛛的背上一段時間,這種親代照顧行為在蜘蛛中相對罕見。狼蛛常見於草地、落葉層和農田等地面環境中。
跳蛛科(Salticidae)是另一個在台灣廣泛分布的蜘蛛科。這類蜘蛛以其敏捷的跳躍能力和發達的視覺系統而聞名。跳蛛的眼睛排列獨特,前端的一對主眼特別大,使它們能夠精確地判斷距離和方向。這些蜘蛛不織網捕獵,而是通過跳躍來捕捉獵物。跳蛛科的許多物種具有鮮豔的體色和複雜的求偶行為,常見於陽光充足的環境中,如樹幹、牆壁或葉面上。
在台灣的室內環境中,我們經常可以看到大魔鬼蛛屬(Heteropoda)的成員,如白額高腳蛛(Heteropoda venatoria)。這種蜘蛛屬於遊蛛科(Sparassidae),體型較大,腿長而有力,能夠快速移動。大魔鬼蛛不織網,而是主動獵捕獵物。它們通常在夜間活動,白天則躲藏在黑暗的角落或縫隙中。雖然外表可能令人生畏,但這種蜘蛛實際上對人類無害,反而能夠幫助控制家中的昆蟲數量。
拱蛛科(Uloboridae)的代表如台灣拱蛛(Cyrtophora moluccensis)也是台灣常見的蜘蛛之一。這種蜘蛛以其獨特的三維網結構而聞名,網呈帳篷狀,通常建在灌木或低矮樹木之間。拱蛛不具有毒腺,而是依靠強韌的絲線來纏繞獵物。它們常形成大型的群居網,多個個體的網彼此相連,形成壯觀的網群。
蟹蛛科(Thomisidae)的成員在台灣的花卉和葉片上也很常見。這類蜘蛛通常體型較小,外形酷似螃蟹,前兩對腳特別發達。蟹蛛擅長偽裝,常常與其棲息的花朵顏色相近,以便於捕捉前來訪花的昆蟲。它們不織網,而是採取埋伏的捕獵策略。
最後,值得一提的是人面蛛科(Theridiosomatidae)。這個科的蜘蛛雖然體型微小,但織造的網卻相當精巧。它們的網通常呈圓錐形,蜘蛛會藏在網的頂端,並通過拉緊中心的絲線來增加網的張力。當獵物接觸到網時,蜘蛛會迅速鬆開絲線,使網收縮並將獵物包裹起來。這種獨特的捕獵方式使人面蛛科在台灣的蜘蛛群落中占據了特殊的生態位。
這些不同科別的蜘蛛展現了台灣蜘蛛fauna的多樣性,從它們的外觀特徵、棲息地選擇到獨特的生活習性,都反映了蜘蛛在長期演化過程中對不同生態環境的適應。這種多樣性不僅豐富了台灣的生態系統,也為蜘蛛學研究提供了寶貴的素材。
3.2.1 白額高腳蛛 (Heteropoda venatoria)
白額高腳蛛(Heteropoda venatoria)是台灣常見的大型蜘蛛之一,屬於遊蛛科(Sparassidae)。這種蜘蛛以其獨特的外觀特徵和生活習性在台灣的蜘蛛fauna中佔有重要地位。
在外觀上,白額高腳蛛體型相當可觀,成年雌蛛體長可達3至4公分,加上腿展可超過12公分。它們的體色通常呈淺褐色或灰褐色,背甲上有深色斑紋,腹部則有不規則的斑點。這種配色使它們能夠很好地融入環境中。大魔鬼蛛最顯著的特徵是其八隻腿又長又有力,使它能夠快速移動和敏捷地捕捉獵物。它們的眼睛排列獨特,共有八隻眼睛分布在頭部前方,形成兩排,這種眼睛的排列有助於它們在夜間進行精確的獵捕活動。
白額高腳蛛的棲息地選擇相當廣泛,但它們特別喜歡人類居住環境的周邊。在都市地區,我們常能在房屋內部、倉庫、車庫等處發現它們的蹤跡。在自然環境中,它們則偏好樹皮下、巖石縫隙或落葉堆等隱蔽的地方。這種棲息地選擇反映了大魔鬼蛛對遮蔽和黑暗環境的偏好,這不僅為它們提供了安全的庇護所,也有利於它們的夜間捕獵活動。
在生活習性方面,白額高腳蛛是一種夜行性捕獵者。與許多蜘蛛不同,它們不織網捕獵,而是主動搜尋和追捕獵物。白天,它們通常躲在隱蔽處休息,到了夜晚則會出來活動和覓食。大魔鬼蛛的食性相當廣泛,包括各種昆蟲、小型蜥蜴,甚至其他蜘蛛。它們敏捷的身手和發達的視覺系統使其成為高效的捕獵者。
白額高腳蛛的繁殖行為也頗為有趣。在求偶過程中,雄蛛會小心翼翼地接近體型較大的雌蛛,並進行一系列複雜的求偶舞蹈。這種行為不僅是為了吸引雌蛛的注意,也是為了避免被誤認為獵物而遭到攻擊。交配成功後,雌蛛會產下數百粒卵並編織成一個扁平的卵囊。與許多蜘蛛不同,雌性大魔鬼蛛會將卵囊緊貼在腹部下方攜帶,直到幼蛛孵化。這種親代照顧行為大大提高了後代的存活率。
值得一提的是,儘管白額高腳蛛的體型和外表可能令人生畏,但它們對人類實際上是無害的。相反,它們在控制家庭和農業害蟲方面發揮著重要作用。許多人認為有大魔鬼蛛出沒的房屋較少受到蟑螂等害蟲的困擾。
白額高腳蛛的存在也引發了一些有趣的文化現象。在台灣的民間傳說中,這種蜘蛛被視為帶來好運的象徵。有些人甚至認為,如果家中有大魔鬼蛛出現,可能預示著即將有財運降臨。這種文化觀念在某種程度上有助於減少人們對這種大型蜘蛛的恐懼,促進了人與自然的和諧共處。
3.2.2 摩鹿加雲斑蛛(Cyrtophora moluccensis)
摩鹿加雲斑蛛(Cyrtophora moluccensis)是台灣常見的一種蜘蛛,屬於拱蛛科(Uloboridae)。這種蜘蛛以其獨特的外觀和織網方式在台灣的蜘蛛群落中佔有特殊地位。
在外觀上,台灣拱蛛呈現出顯著的性二型性。雌蛛體型較大,體長可達15至20毫米,而雄蛛則明顯小得多,通常只有雌蛛的四分之一大小。雌蛛的外觀相當引人注目,背甲呈褐色或灰褐色,腹部則呈現橢圓形,上面有複雜的花紋和突起,這些突起使其看起來像是樹皮或枯葉的一部分,有助於偽裝。雄蛛的體色通常較為單調,呈淺褐色或灰色。
台灣拱蛛最為人知的特徵是其獨特的織網方式。它們織造的網呈三維立體結構,通常被稱為「帳篷網」或「拱形網」。這種網的結構相當複雜,由多層水平絲線和垂直絲線交織而成,形成一個類似帳篷或圓錐形的立體結構。網的頂部較為平坦,而底部則呈漏斗狀。蜘蛛通常棲息在網的上部,背部朝下,這種姿勢有助於它們迅速感知並捕捉落入網中的獵物。
在棲息地選擇方面,台灣拱蛛偏好開闊且有適當支撐點的環境。它們常見於灌木叢、低矮樹木之間,以及建築物的外牆或屋簷下。這些地方提供了足夠的空間讓它們建造大型的立體網。有趣的是,台灣拱蛛經常形成大規模的群居網群,多個個體的網彼此相連,形成一片龐大的網絡。這種群居行為不僅增加了捕獲獵物的效率,也為整個群體提供了更好的防禦能力。
台灣拱蛛的繁殖行為也頗為特別。雄蛛在求偶時會小心翼翼地接近雌蛛的網,並在網的邊緣進行特定的震動,以表明自己的存在。如果雌蛛接受,雄蛛會進入雌蛛的網中進行交配。交配後,雌蛛會在網的上部產下卵囊。卵囊呈扁平狀,通常被絲線牢固地固定在網上。雌蛛會持續守護卵囊直到幼蛛孵化。
在生活習性方面,台灣拱蛛主要以各種飛行昆蟲為食,如蚊子、蛾子和小型飛蟲。它們的網具有極強的黏性,能夠有效捕獲飛行中的昆蟲。當獵物落入網中時,蜘蛛會迅速接近並用絲線將其包裹,然後注入消化酶將獵物液化後吸食。值得注意的是,與許多其他蜘蛛不同,拱蛛科的蜘蛛不具有毒腺,它們完全依靠絲線的力量來制服獵物。
台灣拱蛛的另一個有趣特點是它們的網維護行為。它們會定期清理網上的碎屑,修補破損的部分,並在必要時重建整個網結構。這種持續的維護確保了網的捕獲效率,同時也反映了拱蛛對其棲息環境的適應能力。
在生態系統中,台灣拱蛛扮演著重要的角色。它們不僅是許多飛行昆蟲的天敵,控制這些昆蟲的數量,而且其龐大的網結構也為其他小型生物提供了棲息場所。例如,一些小型寄生蜂和寄生蠅常常在拱蛛的網絡中活動,尋找寄主或獵物。
台灣拱蛛的存在對於研究蜘蛛的社會行為和生態適應具有重要意義。它們的群居行為、複雜的網絡結構以及與周圍環境的互動,為科學家提供了豐富的研究素材。通過研究這種蜘蛛,我們可以更好地理解生物如何適應環境、如何在生態系統中發揮作用,以及群體行為如何影響物種的生存和繁衍。
3.2.3 人面蜘蛛 (Nephila pilipes)
人面蜘蛛(Nephila pilipes)是台灣最為人知的蜘蛛之一,屬於金蛛科(Nephilidae)。這種蜘蛛以其巨大的體型和金黃色的絲線而聞名,在台灣的生態系統中扮演著重要角色。
人面蜘蛛的外觀特徵極為引人注目。雌性金線蛛體型巨大,體長可達4至5公分,若加上腿展則可超過15公分,是台灣最大的蜘蛛之一。雌蛛的腹部呈橢圓形,背面有黃色、白色和黑色相間的斑紋,腹部兩側則有一對明顯的黃色條紋。其八隻腿修長有力,呈現黑色和黃色相間的條紋狀。相比之下,雄性金線蛛的體型要小得多,通常只有雌蛛的十分之一大小,體色也較為單調,呈褐色或灰褐色。這種顯著的性二型性是金線蛛的一大特徵。
金線蛛的名字來源於其織造的金黃色蛛絲。這種絲不僅色澤獨特,而且強度和韌性都極高,能夠承受較大的拉力而不斷裂。金線蛛織造的網可以非常巨大,直徑常常超過一米,有時甚至可達兩米。網呈圓形,由放射狀的支撐絲和螺旋狀的捕獲絲組成。這種大型的圓網通常建在開闊的空間中,如林間空地、庭園或建築物之間。
在棲息地選擇上,人面蜘蛛偏好溫暖潮濕的環境。它們常見於低海拔的森林邊緣、果園、公園和城市綠地等地方。金線蛛需要足夠的空間來建造其巨大的網,因此它們通常選擇樹木或建築物之間的開闊區域。這些區域不僅提供了織網所需的支撐點,也有利於捕捉飛行昆蟲。
金線蛛的繁殖行為非常有趣。由於雌雄體型差異巨大,雄蛛在求偶時需要格外小心。雄蛛會在雌蛛網的邊緣徘徊,等待適當的時機接近雌蛛。有時,多隻雄蛛會聚集在同一張網上,形成激烈的競爭。交配過程中,雄蛛有被雌蛛捕食的風險,因此它們通常會選擇在雌蛛剛蛻皮後或正在進食時接近,以降低被捕食的可能性。交配後,雌蛛會在網附近產下數百粒卵,並用金黃色的絲將其包裹成卵囊。
人面蜘蛛的生活習性主要圍繞其捕獵活動展開。它們主要以各種飛行昆蟲為食,如蝴蝶、蛾子、蜜蜂等。金線蛛的網不僅大,而且黏性極強,能夠有效捕獲體型較大的飛行昆蟲。當獵物落入網中時,金線蛛會迅速接近,用絲線將獵物纏繞,然後注入毒液將其麻痺。有趣的是,金線蛛會根據獵物的大小和類型來調整其捕獲策略。對於較小的獵物,它們可能直接用口器將其咬住;而對於較大或具有危險性的獵物,則會先用大量絲線將其牢牢捆綁。
金線蛛在生態系統中扮演著重要角色。作為一種大型捕食者,它們能夠有效控制許多昆蟲的數量,從而維持生態平衡。此外,金線蛛的網也為其他小型生物提供了棲息場所。例如,有些小型蜘蛛物種會在金線蛛網的邊緣築巢,分享金線蛛捕獲的獵物。
人面蜘蛛的絲線不僅在自然界中發揮作用,也引起了科學界的關注。研究人員發現,金線蛛的絲具有極高的強度和韌性,甚至超過了同重量的鋼絲。這種獨特的特性使得金線蛛絲成為生物材料研究的熱點,科學家們正在探索將其應用於醫療、紡織等領域的可能性。
在台灣的民間文化中,金線蛛也佔有一席之地。許多人認為金線蛛的出現是好運的象徵,特別是當它們在家門口織網時。這種文化觀念在某種程度上有助於保護這種獨特的蜘蛛物種。
3.2.4 長圓金蛛 (Argiope aemula)
長圓金蛛(Argiope aemula)是台灣常見的一種蜘蛛,屬於園蛛科(Araneidae)。這種蜘蛛以其獨特的外觀和織網方式在台灣的蜘蛛群落中佔有重要地位。
在外觀上,長圓金蛛呈現出顯著的性二型性。雌蛛體型較大,體長可達15至25毫米,而雄蛛則明顯小得多,通常只有雌蛛的三分之一大小。雌蛛的外觀相當引人注目,背甲呈銀白色,腹部則呈現橢圓形,上面有黑色、黃色和白色相間的複雜條紋圖案。這些鮮艷的顏色和圖案不僅使其成為台灣最美麗的蜘蛛之一,也可能起到警告潛在捕食者的作用。雄蛛的體色相對較為單調,通常呈褐色或灰色。
長圓金蛛最為人知的特徵是其獨特的織網方式。它們織造的網呈典型的圓形結構,直徑可達50厘米。網的中心區域有一個獨特的之字形或X形的加固結構,稱為「穩定帶」或「裝飾帶」。這種結構的確切功能一直是研究者爭論的話題,有人認為它可以增強網的穩定性,也有人認為它可以吸引獵物或警告鳥類避免撞入蛛網。無論如何,這種獨特的網結構使得伊蚊蛛的網在視覺上非常引人注目。
在棲息地選擇方面,長圓金蛛偏好開闊且有適當支撐點的環境。它們常見於庭園、草地、農田邊緣,以及低矮灌木叢之間。這些地方提供了足夠的空間讓它們建造大型的圓網,同時也有充足的飛行昆蟲作為獵物。伊蚊蛛通常選擇在兩個固定點之間織網,如兩棵灌木之間或草叢與建築物之間。
長圓金蛛的繁殖行為頗為有趣。雄蛛在求偶時會小心翼翼地接近雌蛛的網,並在網的邊緣進行特定的震動,以表明自己的存在。如果雌蛛接受,雄蛛會進入雌蛛的網中進行交配。然而,交配過程對雄蛛來說是極為危險的,因為雌蛛可能會將其當作獵物捕食。為了避免這種情況,雄蛛通常會選擇在雌蛛正在蛻皮或進食時接近。交配後,雌蛛會在網附近產下數百粒卵,並用絲線將其包裹成一個圓形或梨形的卵囊。卵囊通常被懸掛在網的附近,雌蛛會守護卵囊直到幼蛛孵化。
在生活習性方面,長圓金蛛主要以各種飛行昆蟲為食,如蚊子、蠅、蛾等。它們的網具有極強的黏性,能夠有效捕獲飛行中的昆蟲。當獵物落入網中時,蜘蛛會迅速接近並用絲線將其包裹,然後注入毒液將獵物麻痺。伊蚊蛛通常在網的中心區域倒掛等待獵物,這種姿勢使它們能夠迅速感知並捕捉落入網中的獵物。
長圓金蛛還有一個有趣的行為特點,那就是「震網」行為。當感到威脅時,伊蚊蛛會快速地震動身體,使整個網產生劇烈振動。這種行為可能有多重功能:它可以使蜘蛛在視覺上變得模糊,難以被捕食者鎖定;也可能是為了嚇退潛在的威脅;同時,這種振動還可能有助於清理網上的碎屑。
在生態系統中,長圓金蛛扮演著重要的角色。作為一種中型捕食者,它們能夠有效控制許多飛行昆蟲的數量,其中包括一些農業害蟲和傳播疾病的昆蟲如蚊子。因此,伊蚊蛛在自然的病蟲害控制中發揮著重要作用。
長圓金蛛的存在對於研究蜘蛛的行為生態和進化適應具有重要意義。它們鮮艷的體色、獨特的網結構以及複雜的繁殖行為,都為科學家提供了豐富的研究素材。通過研究這種蜘蛛,我們可以更好地理解生物如何通過形態和行為適應來應對環境挑戰,以及性選擇如何影響物種的演化。
在台灣的民間文化中,伊蚊蛛因其美麗的外觀和獨特的網結構而受到人們的喜愛。許多人認為在家門口或庭院中看到伊蚊蛛織網是好運的象徵。這種文化觀念在某種程度上有助於保護這種美麗的蜘蛛物種,促進了人與自然的和諧共處。
3.2.5 縱條銀鱗蛛 (Leucauge magnifica)
縱條銀鱗蛛 (Leucauge magnifica)是台灣常見的一種蜘蛛,屬於長腳蛛科(Tetragnathidae)。這種蜘蛛以其獨特的外觀和織網方式在台灣的蜘蛛群落中佔有特殊地位。
在外觀上,縱條銀鱗蛛 呈現出令人驚嘆的美麗色彩。雌蛛體長約10至15毫米,雄蛛稍小,體長約8至10毫米。其最顯著的特徵是腹部呈現出金屬光澤的銀綠色,上面點綴著黑色、紅色和金色的斑點,形成複雜而艷麗的圖案。這種色彩搭配使得縱條銀鱗蛛 成為台灣最美麗的蜘蛛之一。頭胸部通常呈淺綠色或黃綠色,八隻細長的腿呈現淺綠色或黃色,關節處有深色環紋。
縱條銀鱗蛛 的名字來源於其織造的網的特點。它們織造的網呈水平或微傾斜的圓形結構,直徑通常在30至50厘米之間。網的特點是中心區域非常緊密,而外圍則相對稀疏。最引人注目的是,蜘蛛常常在網的中心位置上方拉一根垂直的單線,這就是「單線」名稱的由來。這根單線可能用於快速逃離潛在威脅,也可能用於感知網的振動。
在棲息地選擇方面,縱條銀鱗蛛 偏好濕潤的環境。它們常見於低海拔的森林邊緣、灌木叢、溪流旁邊,以及庭園中的潮濕角落。這些地方不僅提供了適合織網的空間,也有充足的小型飛行昆蟲作為獵物。單線階蛛通常選擇在植物枝條之間或其他固定點之間織網,網常常建在離地面一至兩米的高度。
縱條銀鱗蛛 的繁殖行為頗為有趣。雄蛛在求偶時會小心翼翼地接近雌蛛的網,並在網的邊緣進行特定的震動,以表明自己的存在。如果雌蛛接受,雄蛛會進入雌蛛的網中進行交配。交配過程相對安全,雌蛛很少會攻擊或捕食雄蛛。交配後,雌蛛會在網附近選擇一個隱蔽的位置產卵。卵通常被包裹在一個小型的、呈球形的絲質卵囊中,卵囊外層常帶有保護色,使其難以被捕食者發現。
在生活習性方面,縱條銀鱗蛛 主要以小型飛行昆蟲為食,如蚊子、果蠅等。它們的網雖然看起來相對簡單,但捕獲效率很高。當獵物落入網中時,蜘蛛會迅速接近並用絲線將其包裹,然後注入毒液將獵物麻痺。單線階蛛通常在網的中心區域倒掛等待獵物,這種姿勢使它們能夠迅速感知並捕捉落入網中的獵物。
縱條銀鱗蛛 有一個有趣的行為特點,那就是「隱藏行為」。當感到威脅時,它們會迅速沿著網中的單線下降,或者快速逃到網下方的植物葉片背面。這種行為使得它們能夠在短時間內消失在捕食者的視線中,大大提高了生存機會。
在生態系統中,縱條銀鱗蛛 扮演著重要的角色。作為一種中小型捕食者,它們能夠有效控制許多小型飛行昆蟲的數量,其中包括一些可能傳播疾病的昆蟲如蚊子。因此,單線階蛛在維持生態平衡和自然的病蟲害控制中發揮著重要作用。
縱條銀鱗蛛 的存在對於研究蜘蛛的行為生態和進化適應具有重要意義。它們美麗的體色可能是為了警告潛在捕食者,這是一個有趣的研究課題。此外,它們獨特的網結構和隱藏行為也為科學家提供了豐富的研究素材。通過研究這種蜘蛛,我們可以更好地理解生物如何通過形態和行為適應來應對環境挑戰。
在台灣的自然教育中,單線階蛛常常被用作教學示例。它們色彩艷麗,容易觀察,且對人類無害,非常適合用於戶外教學活動。通過觀察單線階蛛,學生們可以學習到蜘蛛的基本形態特徵、生活習性,以及它們在生態系統中的重要作用。
值得一提的是,縱條銀鱗蛛 的美麗外表使其成為自然攝影愛好者的熱門拍攝對象。它們金屬光澤的體表在陽光下閃閃發光,加上精巧的網結構,常常能夠創造出令人驚嘆的畫面。這些照片不僅具有極高的審美價值,也有助於提高公眾對蜘蛛以及更廣泛的生物多樣性的認識和欣賞。
3.2.6 尖尾銀鱗蛛 (Leucauge decorata)
尖尾銀鱗蛛(Leucauge decorata)是台灣常見的一種蜘蛛,屬於長腳蛛科(Tetragnathidae)。這種蜘蛛以其獨特的外觀和織網方式在台灣的蜘蛛群落中佔有特殊地位,其名稱「八星」來源於其腹部的特殊圖案。
在外觀上,尖尾銀鱗蛛呈現出令人印象深刻的色彩和圖案。雌蛛體長約8至12毫米,雄蛛略小,體長約6至8毫米。其最顯著的特徵是腹部呈現出銀白色的底色,上面點綴著黑色、紅色和金色的斑點,形成複雜而獨特的圖案。最引人注目的是腹部背面有八個明顯的黑點,排列成兩行,這就是「八星」名稱的由來。頭胸部通常呈淺綠色或黃綠色,八隻細長的腿呈現淺綠色或黃色,關節處有深色環紋。這種色彩搭配使得尖尾銀鱗蛛成為台灣最容易辨認的蜘蛛之一。
尖尾銀鱗蛛織造的網呈水平或微傾斜的圓形結構,直徑通常在20至40厘米之間。網的特點是中心區域非常緊密,而外圍則相對稀疏。與縱條銀鱗蛛 類似,八星階蛛也常常在網的中心位置上方拉一根垂直的單線。這根單線可能用於快速逃離潛在威脅,也可能用於感知網的振動。
在棲息地選擇方面,尖尾銀鱗蛛偏好濕潤的環境。它們常見於低海拔的森林邊緣、灌木叢、溪流旁邊,以及庭園中的潮濕角落。這些地方不僅提供了適合織網的空間,也有充足的小型飛行昆蟲作為獵物。八星階蛛通常選擇在植物枝條之間或其他固定點之間織網,網常常建在離地面0.5至1.5米的高度。
尖尾銀鱗蛛的繁殖行為頗為有趣。雄蛛在求偶時會小心翼翼地接近雌蛛的網,並在網的邊緣進行特定的震動,以表明自己的存在。如果雌蛛接受,雄蛛會進入雌蛛的網中進行交配。交配過程相對安全,雌蛛很少會攻擊或捕食雄蛛。交配後,雌蛛會在網附近選擇一個隱蔽的位置產卵。卵通常被包裹在一個小型的、呈球形的絲質卵囊中,卵囊外層常帶有保護色,使其難以被捕食者發現。
在生活習性方面,尖尾銀鱗蛛主要以小型飛行昆蟲為食,如蚊子、果蠅等。它們的網雖然看起來相對簡單,但捕獲效率很高。當獵物落入網中時,蜘蛛會迅速接近並用絲線將其包裹,然後注入毒液將獵物麻痺。八星階蛛通常在網的中心區域倒掛等待獵物,這種姿勢使它們能夠迅速感知並捕捉落入網中的獵物。
尖尾銀鱗蛛有一個有趣的行為特點,那就是「擬死行為」。當感到威脅時,它們會迅速放下身體,將腿部蜷縮,呈現出一種僵硬的狀態,看起來就像已經死亡一樣。這種行為可以有效地欺騙一些視覺導向的捕食者,如鳥類。當威脅過去後,蜘蛛會迅速恢復正常活動。
在生態系統中,尖尾銀鱗蛛扮演著重要的角色。作為一種中小型捕食者,它們能夠有效控制許多小型飛行昆蟲的數量,其中包括一些可能傳播疾病的昆蟲如蚊子。因此,八星階蛛在維持生態平衡和自然的病蟲害控制中發揮著重要作用。
尖尾銀鱗蛛的存在對於研究蜘蛛的行為生態和進化適應具有重要意義。它們獨特的體色圖案可能是為了警告潛在捕食者或者是一種保護色,這是一個有趣的研究課題。此外,它們的擬死行為也為科學家提供了豐富的研究素材。通過研究這種蜘蛛,我們可以更好地理解生物如何通過形態和行為適應來應對環境挑戰。
在台灣的自然教育中,八星階蛛常常被用作教學示例。它們色彩艷麗,容易辨認,且對人類無害,非常適合用於戶外教學活動。通過觀察八星階蛛,學生們可以學習到蜘蛛的基本形態特徵、生活習性,以及它們在生態系統中的重要作用。
值得一提的是,尖尾銀鱗蛛的美麗外表和獨特圖案使其成為自然攝影愛好者的熱門拍攝對象。它們銀白色的體表和鮮明的黑點在陽光下形成強烈的對比,加上精巧的網結構,常常能夠創造出令人驚嘆的畫面。這些照片不僅具有極高的審美價值,也有助於提高公眾對蜘蛛以及更廣泛的生物多樣性的認識和欣賞。
3.2.9 人面蜘蛛(Nephila pilipes)
人面蜘蛛(Nephila pilipes),又稱為金絲蛛或金蛛,是台灣常見的大型蜘蛛之一,屬於金蛛科(Nephilidae)。這種蜘蛛以其巨大的體型和金黃色的絲線而聞名,在台灣的生態系統中扮演著重要角色。
在外觀上,人面蜘蛛呈現出顯著的性二型性。雌性個體體型巨大,體長可達4至5公分,若加上腿展則可超過15公分,是台灣最大的蜘蛛之一。雌蛛的腹部呈橢圓形,背面有黃色、白色和黑色相間的複雜斑紋,腹部兩側則有一對明顯的黃色條紋。頭胸部較小,呈深褐色或黑色。八隻腿修長有力,呈現黑色和黃色相間的條紋狀。相比之下,雄性個體的體型要小得多,通常只有雌蛛的十分之一大小,體色也較為單調,呈褐色或灰褐色。
人面蜘蛛的名字來源於其腹部背面的圖案,有人認為這些圖案看起來像是一張人臉。然而,這種解讀可能更多是源於人類的想象力,而非蜘蛛本身的特徵。實際上,這些複雜的斑紋可能起到警告或迷惑捕食者的作用。
人面蜘蛛以其金黃色的絲線而聞名。這種絲不僅色澤獨特,而且強度和韌性都極高,能夠承受較大的拉力而不斷裂。人面蜘蛛織造的網可以非常巨大,直徑常常超過一米,有時甚至可達兩米。網呈圓形,由放射狀的支撐絲和螺旋狀的捕獲絲組成。這種大型的圓網通常建在開闊的空間中,如林間空地、庭園或建築物之間。
在棲息地選擇上,人面蜘蛛偏好溫暖潮濕的環境。它們常見於低海拔的森林邊緣、果園、公園和城市綠地等地方。這些蜘蛛需要足夠的空間來建造其巨大的網,因此它們通常選擇樹木或建築物之間的開闊區域。這些區域不僅提供了織網所需的支撐點,也有利於捕捉飛行昆蟲。
人面蜘蛛的繁殖行為非常有趣。由於雌雄體型差異巨大,雄蛛在求偶時需要格外小心。雄蛛會在雌蛛網的邊緣徘徊,等待適當的時機接近雌蛛。有時,多隻雄蛛會聚集在同一張網上,形成激烈的競爭。交配過程中,雄蛛有被雌蛛捕食的風險,因此它們通常會選擇在雌蛛剛蛻皮後或正在進食時接近,以降低被捕食的可能性。交配後,雌蛛會在網附近產下數百粒卵,並用金黃色的絲將其包裹成卵囊。
在生活習性方面,人面蜘蛛主要以各種飛行昆蟲為食,如蝴蝶、蛾子、蜜蜂等。它們的網不僅大,而且黏性極強,能夠有效捕獲體型較大的飛行昆蟲。當獵物落入網中時,人面蜘蛛會迅速接近,用絲線將獵物纏繞,然後注入毒液將其麻痺。有趣的是,人面蜘蛛會根據獵物的大小和類型來調整其捕獲策略。對於較小的獵物,它們可能直接用口器將其咬住;而對於較大或具有危險性的獵物,則會先用大量絲線將其牢牢捆綁。
人面蜘蛛在生態系統中扮演著重要角色。作為一種大型捕食者,它們能夠有效控制許多昆蟲的數量,從而維持生態平衡。此外,人面蜘蛛的網也為其他小型生物提供了棲息場所。例如,有些小型蜘蛛物種會在人面蜘蛛網的邊緣築巢,分享人面蜘蛛捕獲的獵物。
人面蜘蛛的絲線不僅在自然界中發揮作用,也引起了科學界的關注。研究人員發現,人面蜘蛛的絲具有極高的強度和韌性,甚至超過了同重量的鋼絲。這種獨特的特性使得人面蜘蛛絲成為生物材料研究的熱點,科學家們正在探索將其應用於醫療、紡織等領域的可能性。
在台灣的民間文化中,人面蜘蛛也佔有一席之地。許多人認為人面蜘蛛的出現是好運的象徵,特別是當它們在家門口織網時。這種文化觀念在某種程度上有助於保護這種獨特的蜘蛛物種。然而,也有一些人因為人面蜘蛛的巨大體型而感到恐懼。實際上,儘管體型龐大,人面蜘蛛對人類並無實質危害,其毒性對人體影響輕微。因此,提高公眾對這種蜘蛛的了解和欣賞,對於促進人與自然的和諧共處具有重要意義。
3.3 台灣的長腳蛛科Tetragnathidae
長腳蛛科(Tetragnathidae)是台灣常見的蜘蛛科之一,以其修長的腿部和細長的身體為特徵。在台灣,這個科的蜘蛛種類豐富,適應了多樣的生態環境。
台灣常見的長腳蛛科種類包括縱條銀鱗蛛 (Leucauge magnifica)、尖尾銀鱗蛛(Leucauge decorata)和長腳蛛屬(Tetragnatha)的多個種類。這些蜘蛛在外觀上有一些共同的特徵:它們通常有細長的腹部和修長的腿,頭胸部相對較小。許多種類呈現出金屬光澤的體色,如縱條銀鱗蛛 的銀綠色腹部和尖尾銀鱗蛛的銀白色腹部配以黑點。長腳蛛屬的種類則通常呈現褐色或灰色,能很好地融入環境中。
長腳蛛科的蜘蛛在棲息地選擇上顯示出多樣性,但大多偏好潮濕的環境。縱條銀鱗蛛 和尖尾銀鱗蛛常見於低海拔的森林邊緣、灌木叢和溪流附近。它們喜歡在植物枝條之間或其他固定點之間織網,網常常建在離地面0.5至2米的高度。長腳蛛屬的一些種類則更常見於水邊環境,如溪流、池塘或稻田附近。這些蜘蛛常常將網織在水面上方,有些甚至能在水下活動一段時間。
長腳蛛科的網通常呈水平或微傾斜的圓形結構。縱條銀鱗蛛 和尖尾銀鱗蛛的網直徑通常在20至50厘米之間,網的特點是中心區域非常緊密,而外圍則相對稀疏。這些蜘蛛常常在網的中心位置上方拉一根垂直的單線,用於快速逃離或感知振動。長腳蛛屬的網則相對較小,但結構類似。
在繁殖方面,長腳蛛科的行為模式較為一致。雄蛛在求偶時會小心接近雌蛛的網,並在網的邊緣進行特定的震動,以表明自己的存在。如果雌蛛接受,雄蛛會進入雌蛛的網中進行交配。交配後,雌蛛會在網附近選擇一個隱蔽的位置產卵。卵通常被包裹在一個小型的絲質卵囊中,卵囊外層常帶有保護色。
長腳蛛科的生活習性主要圍繞其捕獵活動展開。它們主要以小型飛行昆蟲為食,如蚊子、果蠅等。當獵物落入網中時,蜘蛛會迅速接近並用絲線將其包裹,然後注入毒液將獵物麻痺。這些蜘蛛通常在網的中心區域倒掛等待獵物,這種姿勢使它們能夠迅速感知並捕捉落入網中的獵物。
長腳蛛科的一個有趣行為是「隱藏行為」。當感到威脅時,它們會迅速沿著網中的單線下降,或者快速逃到網下方的植物葉片背面。長腳蛛屬的一些種類則會將身體緊貼在樹枝或草莖上,伸直前腿和後腿,使自己看起來像是植物的一部分。
在台灣的生態系統中,長腳蛛科的豐富度相當高。它們在多種生態環境中都能找到適合的棲息地,從低地到中海拔山區,從自然森林到人工農田,都有長腳蛛科的蹤跡。這種廣泛分布反映了長腳蛛科強大的適應能力。
長腳蛛科在生態系統中扮演著重要角色。作為中小型捕食者,它們能夠有效控制許多小型飛行昆蟲的數量,其中包括一些可能傳播疾病的昆蟲如蚊子。因此,長腳蛛科在維持生態平衡和自然的病蟲害控制中發揮著重要作用。同時,它們也是許多鳥類、蜥蜴和其他較大型捕食者的食物來源。
在台灣的自然教育中,長腳蛛科的蜘蛛常常被用作教學示例。它們色彩艷麗(如縱條銀鱗蛛 和尖尾銀鱗蛛),容易觀察,且對人類無害,非常適合用於戶外教學活動。通過觀察這些蜘蛛,學生們可以學習到蜘蛛的基本形態特徵、生活習性,以及它們在生態系統中的重要作用。
長腳蛛科的存在對於研究蜘蛛的行為生態和進化適應具有重要意義。它們在不同環境中的適應策略,如長腳蛛屬在水邊環境中的特殊行為,為科學家提供了豐富的研究素材。通過研究這些蜘蛛,我們可以更好地理解生物如何通過形態和行為適應來應對不同的生態環境。
3.5 台灣特有種蜘蛛
台灣擁有豐富多樣的生物資源,其中包括許多特有種蜘蛛。這些只在台灣發現的蜘蛛種類,不僅反映了台灣獨特的地理環境和演化歷程,也為台灣的生物多樣性研究提供了寶貴的資料。
台灣的特有種蜘蛛分布在多個科屬中,其中最為人熟知的是台灣大壁虎蛛(Heteropoda taiwanica)。這種蜘蛛屬於異腳蛛科(Sparassidae),體型較大,體長可達2-3厘米。台灣大壁虎蛛通常棲息在森林環境中,特別是在樹皮或岩石縫隙中。它們具有極強的擬態能力,能夠在樹皮上幾乎完美地隱藏自己。這種蜘蛛的發現不僅豐富了台灣的生物多樣性清單,也為研究蜘蛛的適應性進化提供了重要的案例。
另一個值得關注的台灣特有種是台灣斑蛛(Neoscona taiwanica),屬於圓蛛科(Araneidae)。這種蜘蛛以其獨特的背部花紋而聞名,通常在低海拔到中海拔的森林邊緣或開闊地帶築網。台灣斑蛛的研究對於了解台灣本土蜘蛛的生態位和適應策略具有重要意義。例如,研究人員發現這種蜘蛛的網絡結構與其他相近種類有細微的差異,這可能反映了它們對台灣特定環境的適應。
在跳蛛科(Salticidae)中,台灣也有幾個特有種,如台灣麗跳蛛(Siler taiwanensis)。這種小型跳蛛以其敏捷的動作和優秀的視力而聞名。它們通常棲息在低海拔的灌木叢中,以小型昆蟲為食。台灣麗跳蛛的研究為我們提供了寶貴的資訊,特別是在了解島嶼生態系統中小型捕食者的角色方面。
台灣特有的盲蛛科(Oonopidae)成員也值得一提,如台灣絨盲蛛(Gamasomorpha taiwanica)。這種微小的蜘蛛通常生活在落葉層或土壤中,其發現豐富了我們對台灣土壤生態系統的認識。儘管體型小,但這些蜘蛛在維持土壤生態平衡中可能扮演著重要角色。
在高海拔地區,台灣的特有種蜘蛛也有獨特的代表,如台灣高山蛛(Coelotes taiwanensis),屬於漏斗蛛科(Agelenidae)。這種蜘蛛適應了台灣高山環境的低溫和高濕度條件,為研究蜘蛛對極端環境的適應提供了重要案例。台灣高山蛛的研究還有助於我們了解氣候變化對高山生態系統的潛在影響。
值得注意的是,台灣的特有種蜘蛛並不僅限於陸地生態系統。在淡水環境中,也發現了一些特有種,如台灣水蛛(Dolomedes taiwanensis)。這種蜘蛛能夠在水面上行走,甚至潛入水中捕食小魚和水生昆蟲。台灣水蛛的存在突顯了台灣淡水生態系統的獨特性,同時也為研究蜘蛛如何適應水生環境提供了重要線索。
台灣特有種蜘蛛的研究面臨著諸多挑戰。首先,許多特有種的分布範圍較小,種群數量有限,這使得野外觀察和採集變得困難。其次,一些特有種生活在難以到達的環境中,如高山地區或深邃的洞穴,這增加了研究的難度。此外,由於全球氣候變化和人類活動的影響,一些特有種蜘蛛的棲息地正面臨威脅,這使得相關研究工作更顯緊迫。
然而,這些挑戰也帶來了機遇。例如,研究人員開發了一些創新的方法來研究這些難以捉摸的物種。使用紅外相機陷阱來觀察夜行性蜘蛛的行為,或者利用環境DNA技術來檢測稀有種的存在,這些方法都極大地推進了台灣特有種蜘蛛的研究。
台灣特有種蜘蛛的研究不僅對生物學有重要意義,也與生態保育密切相關。這些蜘蛛往往是特定生態系統健康的指標。例如,台灣高山蛛的存在可能反映了高山生態系統的完整性,而台灣水蛛的數量則可能指示淡水環境的質量。因此,監測這些特有種的種群動態,可以為台灣的生態保育工作提供重要參考。
此外,台灣特有種蜘蛛的研究還可能帶來意想不到的科技創新。例如,對人面蜘蛛(Nephila pilipes)蛛絲的研究雖然不是針對特有種,但啟發了新型生物材料的開發。類似地,研究台灣特有種蜘蛛的特殊適應性可能為生物技術領域提供新的靈感。
總的來說,台灣特有種蜘蛛代表了島嶼生態系統的獨特性和脆弱性。它們不僅是台灣生物多樣性的重要組成部分,也是研究演化生物學、生態學和保育生物學的寶貴資源。隨著研究方法的進步和公眾意識的提高,我們有望更深入地了解這些獨特的生物,並為它們的保護做出更多努力。
3.6 台灣的外來入侵種蜘蛛
台灣的地理位置和氣候條件使其成為許多外來物種的理想棲息地。隨著全球化進程的加速,外來入侵種蜘蛛也逐漸成為台灣生態系統面臨的一個重要挑戰。這些蜘蛛通過各種途徑進入台灣,如國際貿易、旅遊等,並在新環境中成功建立種群,對本地生態系統造成不同程度的影響。
其中最為人所知的外來入侵種蜘蛛是褐腺病蛛(Loxosceles reclusa),俗稱褐隱蛛。這種蜘蛛原產於北美洲,被認為是通過貨物運輸inadvertently引入台灣的。褐腺病蛛以其毒性而聞名,其毒液含有一種能夠導致組織壞死的物質。雖然這種蜘蛛性情溫和,不主動攻擊人類,但一旦被咬傷,可能會導致嚴重的皮膚損傷。在台灣,褐腺病蛛主要分布在都市地區,特別是倉庫、地下室等陰暗潮濕的環境中。這種蜘蛛的存在不僅對公共衛生構成潛在威脅,也可能對本地蜘蛛種群造成競爭壓力。
另一個值得關注的外來入侵種是黑寡婦蜘蛛(Latrodectus spp.)。雖然黑寡婦蜘蛛在台灣的分布尚未廣泛,但已有零星報告。這種蜘蛛以其強烈的神經毒素而聞名,雖然咬傷案例罕見,但一旦發生,可能導致嚴重的症狀。黑寡婦蜘蛛通常棲息在乾燥的環境中,如木柴堆或廢棄的建築物中。在台灣,它們主要通過國際貨運inadvertently引入,目前主要在港口和工業區周邊有所發現。
外來入侵種蜘蛛對台灣本地生態系統的影響是多方面的。首先,它們可能與本地蜘蛛種競爭食物和棲息地資源。例如,來自澳大利亞的花園蠅虎(Badumna longinqua)在台灣的都市環境中迅速擴張,可能對本地類似生態位的蜘蛛種造成壓力。這種蜘蛛適應性強,能夠在建築物的縫隙中築網,並以各種小型昆蟲為食。
其次,外來入侵種蜘蛛可能改變本地的捕食-被捕食關係。例如,某些外來蜘蛛可能成為本地鳥類的新食物來源,而另一些則可能捕食本地昆蟲,從而影響整個食物網。在台灣北部,研究人員發現了來自東南亞的長腳蛛(Crossopriza lyoni),這種蜘蛛能夠適應多種環境,其引入可能對本地的小型節肢動物群落產生影響。
此外,一些外來入侵種蜘蛛可能帶來公共衛生問題。除了前面提到的褐腺病蛛和黑寡婦蜘蛛,還有一些潛在的問題種類。例如,澳洲漏斗網蜘蛛(Atrax robustus)雖然目前尚未在台灣發現,但由於其強烈的毒性和對溫暖潮濕環境的適應能力,一直是生態學家密切關注的對象。
然而,並非所有外來蜘蛛都會成為問題物種。有些外來蜘蛛可能在新環境中找到生態位而不會對本地生態系統造成顯著影響。例如,某些來自東南亞的跳蛛(Salticidae)家族成員在台灣的都市綠地中建立了種群,但似乎並未對本地物種造成明顯威脅。這些蜘蛛甚至可能在都市生態系統中發揮一定的害蟲控制作用。
為了應對外來入侵種蜘蛛帶來的挑戰,台灣的研究人員和相關部門採取了多項措施。首先是加強邊境檢疫,通過嚴格的貨物檢查來減少外來蜘蛛的inadvertently引入。其次是建立監測網絡,及時發現和報告新的外來種。例如,台灣的一些大學和研究機構建立了公民科學項目,鼓勵公眾參與外來蜘蛛的監測和報告。
在研究方面,科學家們正在深入研究外來入侵種蜘蛛的生態特性和潛在影響。這包括研究它們的繁殖策略、食性、活動模式等。例如,對於褐腺病蛛,研究人員正在調查其在台灣的分布範圍和種群動態,以評估其潛在的生態和公共衛生風險。同時,也在研究本地天敵對這些外來蜘蛛的控制效果,以尋找可能的生物防治方法。
此外,針對已經建立種群的外來入侵種蜘蛛,台灣的生態學家正在探索有效的管理策略。這包括物理清除、化學防治,以及生態系統管理等方法。例如,在一些都市綠地中,通過適當的植被管理來減少適合外來蜘蛛生存的微環境,從而控制其種群擴張。
值得注意的是,外來入侵種蜘蛛的問題也為台灣的生態系統適應性和彈性提供了一個研究窗口。通過觀察本地生態系統如何應對這些新來者,科學家們可以更好地理解生態系統的動態平衡機制。這些研究不僅對台灣的生態保育工作有重要意義,也為全球的入侵生物學研究提供了寶貴的案例。
最後,提高公眾意識也是應對外來入侵種蜘蛛挑戰的重要一環。通過教育活動和媒體報導,讓公眾了解這些外來蜘蛛的特徵、潛在危險以及正確的應對方法。同時,也要避免引起不必要的恐慌,客觀理性地看待這一生態問題。

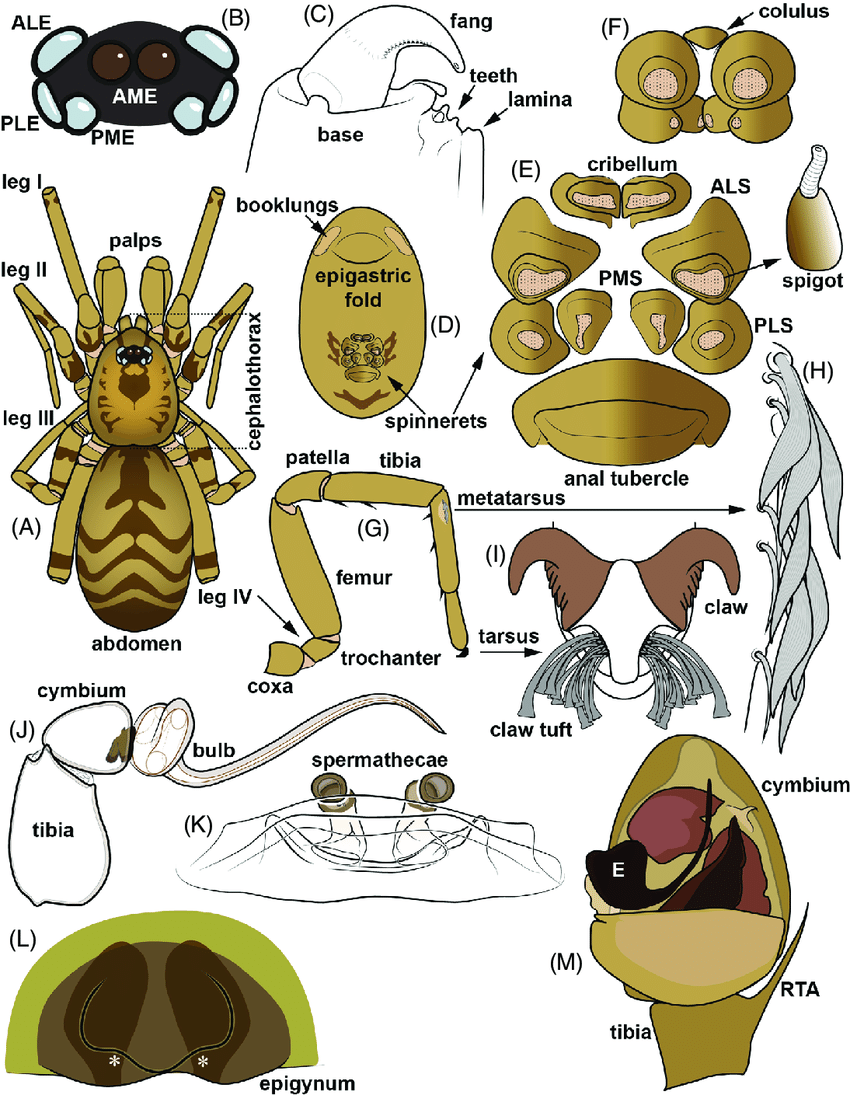
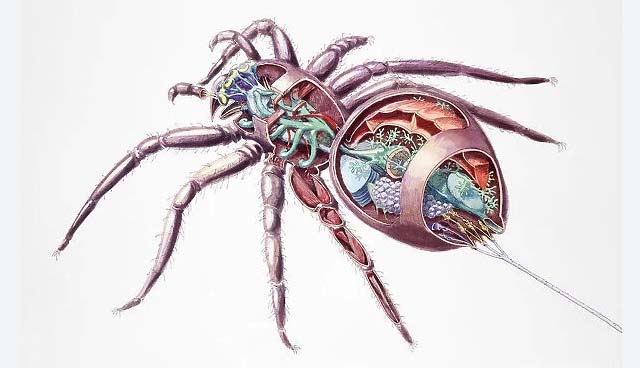
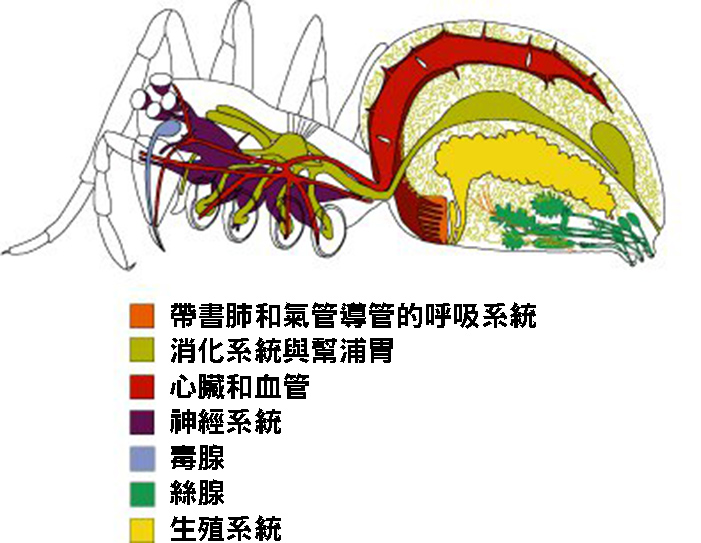
4. 蜘蛛的身體結構
蜘蛛的身體結構是一個精密而複雜的系統,體現了長期演化的結果。蜘蛛屬於節肢動物門蛛形綱,其身體主要分為前體部(prosoma)和後體部(opisthosoma)兩個部分。前體部又稱頭胸部,包含了眼睛、螯肢、觸肢和四對行走腳。後體部則包含了重要的內部器官和絲疣。
蜘蛛的眼睛通常有四對,排列方式因種類而異,這種獨特的眼睛排列有助於它們捕獵和躲避天敵。螯肢是蜘蛛最重要的捕食工具,不僅用於咬住和撕裂獵物,還能分泌毒液。觸肢在雄性蜘蛛中還具有傳遞精子的功能,是一個重要的繁殖器官。
蜘蛛的腹部包含了消化系統、呼吸系統和生殖系統等重要器官。特別值得一提的是位於腹部末端的絲疣,這是蜘蛛產生絲的器官。蜘蛛絲由特殊的蛋白質組成,具有極高的強度和彈性,是蜘蛛捕獵、築巢和移動的重要工具。
蜘蛛獨特的身體結構使它們能夠適應各種環境,從沙漠到熱帶雨林,從高山到洞穴,都能找到蜘蛛的蹤跡。這種適應性強的身體結構也使蜘蛛成為生態系統中重要的捕食者,在維持生態平衡中發揮著關鍵作用。
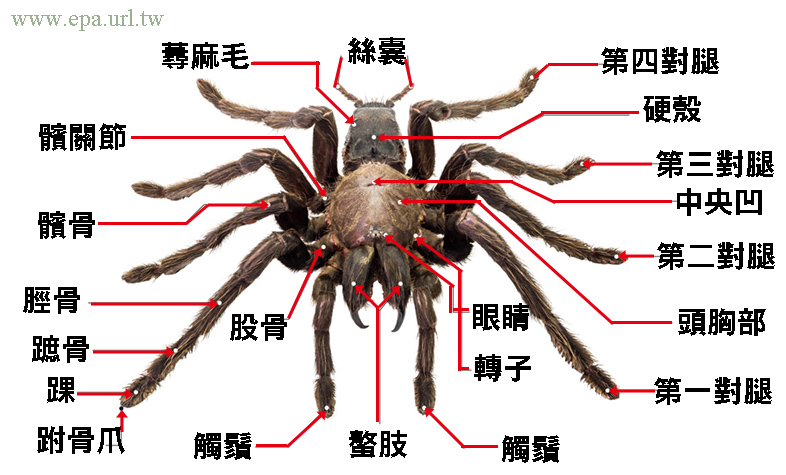
4.1 蜘蛛的外部形態
蜘蛛的外部形態是一個極其精巧且高度專門化的結構,反映了它們在長期演化過程中對各種環境的適應。作為節肢動物門蛛形綱的成員,蜘蛛的身體結構與其他節肢動物有顯著區別,這種獨特的形態使它們能夠在各種生態環境中成功生存。
蜘蛛的身體主要分為兩個部分:前體部(prosoma)和後體部(opisthosoma),這兩個部分通過一個細小的腰部(pedicel)相連。前體部,也稱為頭胸部,是蜘蛛身體的前半部分,包含了許多重要的感覺和運動器官。後體部,通常被稱為腹部,包含了大部分內部器官和特殊的絲腺。
前體部的最顯著特徵是眼睛。大多數蜘蛛擁有八隻眼睛,排列成兩行或三行。這些眼睛的大小、形狀和排列方式在不同種類的蜘蛛中有很大差異,反映了它們的生活方式和捕獵策略。例如,跳蛛科(Salticidae)的蜘蛛擁有極其發達的前中眼,這使它們能夠進行精確的視覺定位,有助於它們捕捉快速移動的獵物。相比之下,織網蜘蛛如圓網蛛(Araneidae)的眼睛相對較小,因為它們主要依賴網絡振動來感知獵物。
在前體部的前端是一對強大的螯肢(chelicerae)。螯肢是蜘蛛最重要的捕食工具,由基節和可動的鉤爪組成。鉤爪的內側有毒腺開口,能夠分泌毒液。螯肢不僅用於捕獲和殺死獵物,還用於撕碎食物,有些蜘蛛甚至用它們來挖掘巢穴。例如,澳洲的漏斗網蛛(Atrax robustus)就有特別發達的螯肢,不僅用於捕食,還用於挖掘複雜的地下巢穴系統。
在螯肢的兩側是一對觸肢(pedipalps)。觸肢在雌性蜘蛛中主要用於感知和操作食物,而在雄性蜘蛛中,觸肢的末端演化成了複雜的交配器官。這種性別二態性在許多蜘蛛種類中非常明顯。例如,在跳蛛中,雄性的觸肢末端膨大,形成了複雜的結構,用於儲存和傳遞精子。
蜘蛛的四對行走腳也附著在前體部。每條腿通常由七節組成,從基部到末端依次是基節、轉節、腿節、膝節、脛節、跗節和爪節。腿的長度和形態因種類而異,反映了蜘蛛的生活習性。例如,攀爬蛛(Crab spiders)的腿相對較短,有利於它們在花朵和樹葉上隱藏和捕獵;而長腳蛛(Pholcidae)的腿則非常細長,有助於它們在複雜的網絡結構中移動。
蜘蛛的後體部,也就是腹部,通常呈橢圓形或球形,但在不同種類中可能有很大變化。例如,盾蛛科(Thomisidae)的某些成員腹部呈扁平狀,有助於它們在樹皮縫隙中隱藏。腹部的背面通常有各種顏色和圖案,這些不僅有助於蜘蛛隱藏或警告天敵,還可能在求偶過程中起作用。
腹部最顯著的外部特徵是位於末端的絲疣(spinnerets)。大多數蜘蛛有三對絲疣,但數量可能因種類而異。絲疣是特化的附屬物,內部連接著絲腺,用於產生和操控蛛絲。不同的絲疣可以產生不同類型的絲,用於不同的目的,如構建捕獵網、包裹獵物、製作蛋囊等。
在某些蜘蛛種類中,腹部還可能有其他特殊結構。例如,某些圓網蛛的腹部上有明顯的突起或棘刺,這可能有助於它們在植物中偽裝。另一個例子是板腹蛛科(Theridiidae)的某些成員,它們的腹部呈三角形,有助於它們在不規則網中隱藏。
蜘蛛的外部形態還包括許多微小但重要的結構。例如,在腿部和身體其他部位分布著各種感覺毛和感受器,這些結構能夠感知空氣振動、化學信號和觸覺刺激,對蜘蛛的生存至關重要。某些蜘蛛,如狼蛛科(Lycosidae)的成員,在腿部還有特殊的梳狀毛,用於感知地面振動。
蜘蛛的外部形態不僅反映了它們的生活方式和生態適應,也為我們研究生物演化提供了豐富的材料。通過比較不同蜘蛛種類的外部形態,科學家們可以推斷它們的親緣關係和演化歷史。例如,原蛛亞目(Mesothelae)的成員保留了許多原始特徵,如分節的腹部,這為我們理解蜘蛛的演化提供了重要線索。
此外,蜘蛛的外部形態還為生物仿生學提供了豐富的靈感。例如,蜘蛛腳部的特殊結構啟發了科學家開發新型的粘附材料,而蜘蛛絲的獨特性質則促進了高強度纖維材料的研究。這些研究不僅加深了我們對蜘蛛生物學的理解,也為人類科技的發展做出了貢獻。

4.1.1 蜘蛛的外觀色彩圖案與絨毛
蜘蛛的外觀色彩圖案與絨毛是其外部形態中最引人注目的特徵之一,不僅體現了蜘蛛的多樣性,也反映了它們在長期演化過程中對環境的適應。這些外觀特徵在蜘蛛的生存、捕食和繁衍中扮演著關鍵角色,同時也為我們研究生物適應性和演化提供了豐富的素材。
蜘蛛的體色範圍極其廣泛,從單調的褐色、灰色到鮮艷的紅色、黃色、綠色等,甚至有些蜘蛛還具有金屬光澤。這種多樣的色彩通常與蜘蛛的生活環境密切相關。例如,生活在樹皮上的蜘蛛往往呈現褐色或灰色,有助於它們在背景中隱藏。而生活在花朵上的蜘蛛,如某些盾蛛科(Thomisidae)的成員,則可能呈現鮮艷的黃色或粉色,以便在花中偽裝。
蜘蛛的體色還可能隨環境變化而改變。一個著名的例子是螃蟹蜘蛛(Misumena vatia),它能夠根據所處的花朵顏色調整自身的體色,從白色變為黃色,或反之,這種能力使它能夠更好地隱藏自己並捕捉訪花昆蟲。
除了整體色彩,許多蜘蛛還具有複雜的體表圖案。這些圖案可能包括條紋、斑點、環帶或幾何形狀等。例如,花園蜘蛛(Araneus diadematus)背部的十字形圖案,或是黑寡婦蜘蛛(Latrodectus spp.)腹部的紅色沙漏形標記。這些圖案可能具有多種功能,包括偽裝、警戒色或作為種間識別的標誌。
某些蜘蛛的體表圖案還可能隨年齡或性別而變化。例如,在許多跳蛛科(Salticidae)的物種中,雄性often比雌性有更鮮艷的色彩和更複雜的圖案,這在求偶過程中起著重要作用。雄性跳蛛會通過展示其色彩鮮艷的部位來吸引雌性的注意。
蜘蛛的絨毛是另一個重要的外觀特徵。這些細小的毛髮覆蓋在蜘蛛的體表,不僅賦予了蜘蛛獨特的質地,還具有多種重要功能。首先,絨毛可以增強蜘蛛的偽裝效果。例如,某些地蜘蛛科(Gnaphosidae)的成員體表覆蓋著灰色或褐色的絨毛,使它們能夠完美地融入土壤或樹皮背景中。
其次,絨毛在蜘蛛的感知系統中扮演著重要角色。許多絨毛實際上是特化的感覺器官,能夠感知空氣振動、化學信號和觸覺刺激。例如,狼蛛科(Lycosidae)的成員在腿部有特殊的絨毛,可以感知地面振動,幫助它們定位獵物或識別潛在威脅。
某些蜘蛛的絨毛還具有特殊的光學性質。例如,跳蛛科中的孔雀蜘蛛(Maratus spp.),雄性的腹部覆蓋著能夠反射不同顏色光線的鱗片狀絨毛,產生類似孔雀尾羽的炫麗效果。這種絢爛的"彩衣"在求偶過程中起著關鍵作用。
蜘蛛的絨毛還可能具有防禦功能。某些蜘蛛,如鳥蛛科(Theraphosidae)的成員,體表覆蓋著刺激性的剛毛。當受到威脅時,這些蜘蛛會用後腿快速摩擦腹部,將這些刺激性的毛髮射向潛在的捕食者,造成強烈的刺激和不適。
蜘蛛的外觀色彩圖案和絨毛不僅反映了它們的生活環境和行為特徵,還為我們研究生物適應性和演化提供了豐富的素材。例如,通過比較不同環境中同種蜘蛛的色彩變異,科學家們可以研究環境對生物特徵的選擇壓力。同時,蜘蛛的外觀特徵也為生物仿生學提供了靈感。例如,某些蜘蛛的結構色啟發了新型光學材料的開發。
值得注意的是,蜘蛛的外觀特徵可能會隨季節、年齡或生理狀態而變化。例如,某些蜘蛛在冬季可能會變得更暗,以吸收更多的熱量。而在蛻皮過程中,蜘蛛的外觀也會暫時改變,新的外骨骼often需要一段時間才能完全硬化和著色。
此外,蜘蛛的外觀特徵還可能受到寄生蟲或疾病的影響。例如,某些被真菌感染的蜘蛛可能會出現異常的白色斑點。了解這些變化不僅對研究蜘蛛的健康狀況很重要,也有助於我們理解更廣泛的生態系統動態。
在研究蜘蛛的外觀特徵時,科學家們常常使用顯微鏡和特殊的攝影技術。例如,掃描電子顯微鏡可以揭示蜘蛛絨毛的微觀結構,而紫外光攝影則可以顯示出人眼無法看到的紫外反射圖案。這些技術幫助我們更深入地理解蜘蛛外觀特徵的功能和演化意義。
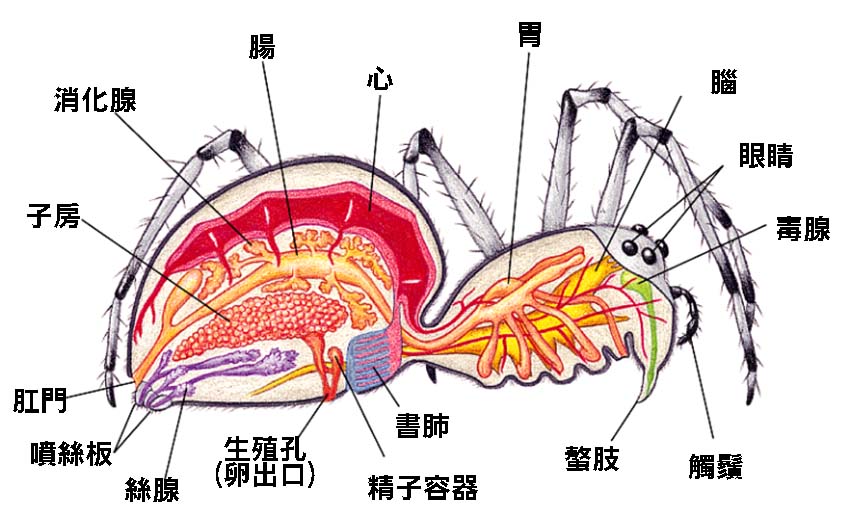
4.2 蜘蛛的內部器官
蜘蛛的內部器官系統是一個精密而高效的結構,反映了它們在長期演化過程中對環境的適應。儘管蜘蛛的身體相對較小,但其內部器官卻十分複雜,能夠支持它們的各種生理功能和行為模式。
蜘蛛的消化系統是其內部器官中最顯著的部分之一。這個系統始於口器,包括一對螯肢和一對觸肢。螯肢不僅用於捕獲和撕碎獵物,還連接著毒腺,可以分泌毒液。毒液不僅用於麻痺獵物,還具有消化功能,能夠在獵物體內開始初步的消化過程。
從口器開始,食物進入前腸,這裡包括咽、食道和胃。蜘蛛的胃結構特殊,能夠產生強大的吸力,將液化的食物吸入中腸。中腸是蜘蛛消化系統的核心部分,由多個分支組成,延伸到腹部和腿部。這種結構增加了消化和吸收的表面積,使蜘蛛能夠高效地利用食物。例如,跳蛛科(Salticidae)的成員,儘管體型小,但其中腸分支發達,能夠快速消化獵物,支持它們活躍的捕獵行為。
蜘蛛的循環系統是開放式的,由心臟和血竅組成。心臟位於腹部背側,呈管狀,具有多對側孔。血液(血淋巴)通過這些側孔進入心臟,然後被泵送到全身。這種開放式循環系統雖然看似簡單,但對於蜘蛛這樣的小型節肢動物來說非常有效。例如,狼蛛科(Lycosidae)的成員具有發達的心臟肌肉,能夠支持它們快速奔跑和跳躍的行為。
蜘蛛的呼吸系統主要由書肺和氣管組成。書肺是蜘蛛特有的呼吸器官,位於腹部腹面,由多層薄膜疊加而成,形如書頁,因此得名。大多數蜘蛛有一對或兩對書肺。某些蜘蛛,如跳蛛和狼蛛,還具有氣管系統,這使它們能夠更有效地供氧,支持高度活躍的生活方式。
蜘蛛的排泄系統相對簡單,主要由馬氏管和肛門組成。馬氏管是蜘蛛的主要排泄器官,負責過濾血淋巴中的代謝廢物。這些廢物最終通過肛門排出體外。有趣的是,某些蜘蛛,如金蛛科(Nephilidae)的成員,能夠重吸收馬氏管中的水分,這使它們能夠在乾旱環境中生存。
蜘蛛的神經系統高度集中,主要由圍食道神經節和腹神經節組成。圍食道神經節相當於蜘蛛的"大腦",控制著感覺和運動功能。腹神經節則控制腹部和後肢的活動。這種集中的神經系統使蜘蛛能夠快速處理信息和做出反應,這在捕獵和躲避天敵時尤為重要。例如,跳蛛的中央神經系統特別發達,支持其複雜的視覺處理和精確的跳躍行為。
蜘蛛的生殖系統位於腹部,雌雄分開。雄性蜘蛛的生殖系統包括一對精巢、輸精管和儲精囊。有趣的是,雄性蜘蛛使用變形的觸肢(交接器)來傳遞精子,這是蜘蛛獨特的繁殖適應。雌性蜘蛛的生殖系統包括卵巢、輸卵管和交配囊。某些蜘蛛,如黑寡婦(Latrodectus spp.),具有複雜的交配囊結構,能夠儲存多個雄性的精子。
蜘蛛的絲腺是另一個獨特而重要的內部器官。絲腺位於腹部,通過絲疣分泌蛛絲。不同種類的蜘蛛可能有不同數量和類型的絲腺,產生不同性質的絲。例如,金蛛科的成員擁有特別發達的絲腺,能夠產生極其堅韌的絲,用於構建大型捕獵網。
蜘蛛還擁有特殊的內分泌系統,包括各種激素分泌腺。這些腺體調控著蜘蛛的生長、發育、蛻皮和生殖等過程。例如,蛻皮激素控制著蜘蛛的周期性蛻皮,這對蜘蛛的生長至關重要。
值得注意的是,蜘蛛的某些內部器官具有多功能性。例如,肝胰腺不僅參與消化,還在解毒和儲存營養物質方面發揮作用。這種多功能性使蜘蛛能夠在有限的體內空間中實現複雜的生理功能。
蜘蛛的內部器官還表現出顯著的適應性。例如,沙漠蜘蛛的馬氏管特別發達,能夠高效地保存水分。而某些生活在高海拔地區的蜘蛛,如高山漏斗蛛(Coelotes spp.),其書肺結構特殊,能夠在低氧環境中維持有效的氣體交換。
研究蜘蛛的內部器官不僅有助於我們理解這些生物的生理功能,還為生物醫學和材料科學提供了寶貴的靈感。例如,對蜘蛛毒腺的研究促進了新型藥物的開發,而蜘蛛絲腺的研究則推動了高性能生物材料的發展。
4.3 蜘蛛的絲腺和絲疣
蜘蛛的絲腺和絲疣是這些八腳生物最為獨特和引人注目的器官之一,它們不僅是蜘蛛生存的關鍵,也是科學研究和生物技術創新的重要靈感來源。這些精密的器官使蜘蛛能夠製造出令人驚嘆的蛛絲,這種材料以其強度、彈性和多功能性而聞名。
絲腺是位於蜘蛛腹部的特殊腺體,負責產生蛛絲蛋白。每種蜘蛛通常擁有多種不同類型的絲腺,每種絲腺產生特定性質的絲。常見的絲腺類型包括大隆起絲腺、小隆起絲腺、鞭狀絲腺、管狀絲腺、聚合絲腺和梨形絲腺等。這些不同的絲腺產生的絲具有不同的特性和功能,使蜘蛛能夠應對各種需求。
例如,大隆起絲腺產生的絲通常用於構建網的框架或蜘蛛的拖曳線,這種絲具有極高的強度和彈性。小隆起絲腺產生的絲則常用於包裹獵物或製作蛋囊。鞭狀絲腺產生的絲具有很強的黏性,主要用於捕獲獵物。管狀絲腺產生的絲often用於製作蛋囊的外層保護結構。
不同種類的蜘蛛可能擁有不同數量和類型的絲腺。例如,織網蜘蛛如圓網蛛(Araneidae)擁有更多種類的絲腺,以滿足其複雜的織網需求。而一些不織網的蜘蛛,如狼蛛(Lycosidae),可能只有較少種類的絲腺。
絲疣是蜘蛛腹部末端的特化結構,是蛛絲分泌到體外的出口。大多數蜘蛛擁有三對絲疣:前絲疣、中絲疣和後絲疣。每個絲疣上都有數百個微小的開口,稱為紡絲管。不同的絲腺通過不同的紡絲管將絲輸送到體外。
絲疣的結構和功能非常複雜。蜘蛛能夠精確控制每個絲疣的運動,從而決定哪些絲腺分泌絲,以及如何將不同類型的絲組合在一起。這種精確的控制使蜘蛛能夠根據需要製造出不同性質的絲線。例如,當蜘蛛需要製作具有黏性的捕獲絲時,它會同時啟動產生骨架絲的絲腺和產生黏性物質的絲腺,並通過絲疣將兩種物質結合在一起。
某些蜘蛛還擁有特殊的絲疣結構。例如,梳狀蛛(Cribellate spiders)擁有一種稱為梳板(Cribellum)的特殊絲疣,這種結構可以產生極細的絲,用於製造具有特殊捕獲能力的網。
蜘蛛絲的產生過程是一個複雜的生物化學過程。在絲腺中,蛛絲蛋白以液態形式存在。當這種液態蛋白通過紡絲管時,會受到機械力和化學環境的改變,導致蛋白質分子重新排列,形成堅固的絲線。這個過程發生得非常快,幾乎是瞬間完成的。
蜘蛛對其絲腺和絲疣的使用展現出令人驚嘆的靈活性和創造性。例如,某些蜘蛛物種能夠製造出具有特殊功能的絲。跳蛛(Salticidae)就能夠產生一種特殊的「安全線」,在跳躍時防止跌落。某些蜘蛛甚至能夠製造出具有特殊氣味的絲,用於吸引異性。
蜘蛛的絲腺和絲疣不僅在其生活中扮演重要角色,也為科學研究提供了豐富的素材。研究人員對蛛絲的特性和製造過程進行了深入研究,希望能夠開發出類似的人造材料。例如,科學家們已經成功地在轉基因山羊和蠶中產生了蛛絲蛋白,這為大規模生產蛛絲材料開闢了可能性。
此外,蜘蛛絲的研究還為醫學領域帶來了新的機遇。研究人員發現,某些蛛絲蛋白具有促進傷口癒合的潛力,可能用於開發新型的醫療敷料。同時,蛛絲的生物相容性也使其成為潛在的組織工程支架材料。
蜘蛛的絲腺和絲疣還為我們提供了研究生物適應性和演化的寶貴案例。不同種類蜘蛛的絲腺和絲疣結構的差異,反映了它們對不同生態環境的適應。例如,生活在乾旱環境中的蜘蛛往往具有能夠產生更耐久絲線的絲腺,以減少水分損失。
然而,儘管科學家們對蜘蛛的絲腺和絲疣進行了大量研究,但仍有許多未解之謎。例如,蜘蛛如何在分子層面上控制蛛絲的形成過程,以及不同種類蜘蛛的絲腺如何在演化過程中發生分化等問題,仍然是研究的熱點。
4.4 蜘蛛的感覺器官
蜘蛛的感覺器官是一套精密而高效的系統,使這些八腳生物能夠敏銳地感知周圍環境,捕捉獵物,躲避天敵,並與同類交流。這些感覺器官包括視覺、觸覺、化學感受、振動感受等多個方面,每一種都經過長期演化,以適應蜘蛛的特殊生存需求。
視覺是蜘蛛最引人注目的感覺之一。大多數蜘蛛擁有八隻眼睛,但眼睛的大小、排列和功能在不同種類間有顯著差異。這些眼睛通常分為主眼和側眼。主眼通常較大,能夠形成清晰的影像,而側眼主要用於感知光線強度和運動。
跳蛛科(Salticidae)的視覺系統尤其發達,是所有無脊椎動物中最為複雜的之一。它們的前中眼特別大,能夠形成清晰的彩色影像,視力可與小型哺乳動物相媲美。這種優秀的視力使跳蛛能夠精確地定位和捕捉獵物,甚至能夠識別複雜的視覺信號進行求偶。例如,孔雀蜘蛛(Maratus spp.)雄性複雜的求偶舞蹈就依賴於雌性的高度視覰能力。
相比之下,某些蜘蛛的視覺能力相對較弱。例如,許多織網蛛的眼睛較小,主要用於感知光線變化和簡單的運動。這些蜘蛛更多地依賴其他感覺器官,如振動感受器,來感知獵物的存在。
觸覺是蜘蛛另一個重要的感覺系統。蜘蛛的整個身體,特別是腿部,都覆蓋著大量的感覺毛。這些毛髮不僅能感知物理接觸,還能感知氣流變化。例如,某些遊蕩蜘蛛(Wandering spiders)能夠通過感知氣流來判斷飛行昆蟲的位置和大小,從而精確地捕獲獵物。
蜘蛛的化學感受器主要分布在腿部和觸肢上。這些感受器能夠感知空氣中的化學物質,幫助蜘蛛識別食物、天敵和潛在的配偶。例如,雄性蜘蛛能夠通過感知雌性留下的信息素來追蹤配偶。某些蜘蛛,如狼蛛科(Lycosidae)的成員,還能通過化學感受器來識別同類,這在它們的領地行為中起著重要作用。
振動感受是蜘蛛最為依賴的感覺之一,特別是對於織網蛛來說。蜘蛛的腿部和身體上分布著大量的裂縫感受器(Slit sensilla),這些器官能夠感知極其微小的振動。對於織網蛛來說,這種能力使它們能夠精確地定位落入網中的獵物。例如,金園蛛(Nephila spp.)能夠通過網絲的振動來判斷獵物的大小和位置,從而決定是否接近捕獲。
某些蜘蛛還擁有特殊的振動感受器官。例如,漏斗網蛛科(Agelenidae)的成員在腿部擁有被稱為毛狀裂縫感受器(Trichobothria)的特殊結構,這種結構對空氣振動特別敏感,使這些蜘蛛能夠在黑暗的環境中也能精確捕獵。
蜘蛛還可能擁有其他特殊的感覺器官。例如,某些生活在洞穴或地下的蜘蛛可能發展出對溫度和濕度變化敏感的器官。一些水生蜘蛛,如姬蛛科(Cybaeidae)的某些成員,則可能擁有感知水流的特殊感覺器官。
值得注意的是,蜘蛛的感覺系統often是高度整合的。例如,在捕獵過程中,蜘蛛可能同時使用視覺、觸覺和振動感受來定位和捕捉獵物。這種多感官整合使蜘蛛能夠在複雜的環境中高效地生存。
蜘蛛的感覺器官還表現出顯著的適應性。例如,生活在黑暗環境中的蜘蛛可能退化視覺系統,但發展出更敏銳的觸覺和振動感受能力。而生活在明亮開闊環境中的蜘蛛,如跳蛛,則可能擁有更發達的視覺系統。
研究蜘蛛的感覺器官不僅有助於我們理解這些生物的行為和生態,還為生物感測技術的發展提供了寶貴的靈感。例如,科學家們正在研究如何模仿蜘蛛的振動感受系統,以開發更靈敏的地震探測器。蜘蛛的化學感受器也為開發新型氣體傳感器提供了思路。
此外,蜘蛛感覺器官的研究還為神經科學領域帶來了新的見解。例如,跳蛛的視覺系統為研究小型大腦如何處理複雜視覺信息提供了理想的模型。這些研究不僅加深了我們對蜘蛛的理解,也為更廣泛的神經科學和認知科學研究做出了貢獻。
5. 蜘蛛的生理適應
蜘蛛的生理適應是一個複雜而精妙的過程,反映了這些生物在長期演化中對不同環境的適應能力。這些適應涵蓋了多個方面,包括呼吸、循環、代謝和體溫調節等生理功能。
蜘蛛的呼吸系統主要由書肺和氣管組成,能夠高效地進行氣體交換。書肺的結構類似於摺疊的書頁,提供了大面積的氣體交換表面,使蜘蛛能夠在靜止狀態下維持基本的氧氣需求。而氣管系統則能夠在蜘蛛活動時提供額外的氧氣供應。
循環系統方面,蜘蛛擁有開放式循環系統,血液(血淋巴)直接浸潤組織器官。這種系統雖然簡單,但對於體型較小的蜘蛛來說非常有效,能夠快速地將氧氣和營養物質輸送到全身。
蜘蛛還表現出對極端環境的顯著適應能力。例如,某些沙漠蜘蛛能夠長期忍受高溫和缺水環境,這得益於它們特殊的體液調節機制和低代謝率。而生活在寒冷地區的蜘蛛則可能產生類似於防凍劑的物質,防止體內結冰。
此外,蜘蛛的毒液系統也是其重要的生理適應之一。毒液不僅用於捕獵和防禦,還在一定程度上參與消化過程。不同種類的蜘蛛可能進化出不同成分的毒液,以適應特定的獵物或生存環境。
5.1 蜘蛛的呼吸系統
蜘蛛的呼吸系統是一個精巧而高效的結構,體現了這些八腳動物對陸地生活的卓越適應。與其他陸生節肢動物不同,蜘蛛擁有獨特的呼吸器官組合,主要包括書肺和氣管系統,有些物種還保留了原始的腹肺。這種複雜的呼吸系統使蜘蛛能夠在各種環境中生存並保持高度的活動能力。
書肺是蜘蛛最具特色的呼吸器官。顧名思義,書肺的結構類似於一本打開的書,由許多薄薄的組織片層堆疊而成,這些片層之間充滿空氣。書肺通常位於蜘蛛腹部的前端,通過腹部表面的氣孔與外界相通。當蜘蛛吸氣時,空氣進入書肺內部,氧氣通過這些薄層擴散到血液中,同時二氧化碳從血液擴散到空氣中被排出體外。這種結構大大增加了氣體交換的表面積,使得蜘蛛能夠高效地進行呼吸。
不同種類的蜘蛛可能擁有不同數量的書肺。例如,原蛛亞目(Mesothelae)的成員擁有兩對書肺,這被認為是較為原始的特徵。而大多數現代蜘蛛,如園蛛科(Araneidae)的成員,只有一對書肺。有趣的是,某些蜘蛛,如跳蛛科(Salticidae)的物種,完全沒有書肺,而是完全依賴氣管系統呼吸。
氣管系統是蜘蛛呼吸系統的另一個重要組成部分。氣管是一系列細小的管道,從體表的氣孔延伸到體內各個組織。這種系統能夠直接將氧氣輸送到需要的組織,特別適合為活躍的肌肉提供氧氣。擁有發達氣管系統的蜘蛛,如狼蛛科(Lycosidae)的成員,通常具有較強的運動能力和耐力。
某些蜘蛛物種還保留了被稱為腹肺的原始呼吸器官。腹肺的結構較為簡單,基本上是腹部表面的凹陷區域。雖然效率不如書肺,但在某些生活環境中仍然有其優勢。例如,某些生活在潮濕環境中的蜘蛛,如某些洞穴蜘蛛,就保留了腹肺。
蜘蛛的呼吸系統展現出驚人的適應性。例如,某些水生蜘蛛,如水蜘蛛(Argyroneta aquatica),能夠利用特殊的毛髮結構在水下攜帶空氣泡,形成一個臨時的「鐘罩」,使它們能夠在水下長時間呼吸。這種適應使水蜘蛛能夠完全適應水生環境,甚至在水下產卵和養育幼蛛。
高海拔環境中的蜘蛛也表現出獨特的呼吸適應。例如,某些生活在喜馬拉雅山區的蜘蛛物種,其書肺結構有所改變,能夠在低氧環境中更有效地吸收氧氣。這種適應使這些蜘蛛能夠在海拔超過5000米的環境中生存。
蜘蛛的呼吸系統還與其行為和生態習性密切相關。例如,某些蜘蛛在休息時會採取特定的姿勢,以確保書肺能夠充分接觸空氣。織網蜘蛛通常會在網中心保持一定的姿勢,這不僅有利於感知獵物,也能確保最佳的呼吸效率。
值得注意的是,蜘蛛的呼吸系統與其他生理系統緊密聯繫。例如,蜘蛛的開放式循環系統與呼吸系統協同工作,確保氧氣能夠迅速到達全身組織。同時,蜘蛛的排泄系統,特別是馬氏管,也參與調節體內氣體平衡,協助呼吸過程。
蜘蛛的呼吸系統還表現出顯著的可塑性。例如,某些spiders能夠根據環境條件調整其呼吸率。在高溫或運動時,蜘蛛可能會增加呼吸頻率以滿足增加的氧氣需求。相反,在低溫或休眠狀態下,蜘蛛可以顯著降低呼吸率以節省能量。
研究蜘蛛的呼吸系統不僅有助於我們理解這些生物的生理機制,還為生物醫學領域提供了寶貴的靈感。例如,蜘蛛書肺的結構啟發了科學家們在開發新型人工肺和氣體交換膜方面的研究。同時,蜘蛛在極端環境下的呼吸適應也為研究人類在高海拔或其他低氧環境中的生理反應提供了重要參考。
5.2 蜘蛛的循環系統
蜘蛛的循環系統是一個精巧而高效的結構,充分體現了這些節肢動物對陸地生活的適應。與高等脊椎動物不同,蜘蛛擁有開放式循環系統,這種系統雖然結構相對簡單,但對於蜘蛛這樣的小型生物來說卻非常有效。
蜘蛛的循環系統主要由心臟、動脈和血竇組成。心臟是整個系統的核心,它位於蜘蛛腹部的背側,呈管狀結構,通常延伸蜘蛛腹部的大部分長度。心臟的壁上有多對側孔,這些孔在心臟舒張時打開,允許血液(在節肢動物中通常稱為血淋巴)進入心臟;在心臟收縮時關閉,防止血液回流。
心臟的跳動方式非常有趣。它不是像人類心臟那樣從一端到另一端收縮,而是分段收縮。這種收縮方式能夠產生一種波浪式的運動,有效地將血液泵送到身體各部分。例如,在某些大型蜘蛛如狼蛛(Lycosidae)中,研究人員觀察到心臟跳動頻率可以根據活動狀態而變化,在靜止時可能每分鐘跳動40-50次,而在劇烈運動時可以增加到每分鐘200次以上。
從心臟發出的主動脈向前延伸,進入頭胸部,然後分支成幾條較小的動脈,將血液輸送到腿部、眼睛和其他器官。這些動脈在到達組織後開放,血液直接流入組織間隙,形成血竇。血竇是開放式循環系統的一個重要特徵,它們實際上是體腔中的空間,被各種組織和器官包圍。
在血竇中,血液直接與組織接觸,進行氣體、營養物質和代謝產物的交換。這種直接交換的方式雖然效率不如封閉式循環系統高,但對於體型較小的蜘蛛來說已經足夠。而且,這種系統的一個優勢是可以在蜘蛛運動時為肌肉提供額外的液壓支持。
血液完成交換後,通過特定的通道回流到心臟周圍的心包腔。心包腔是圍繞心臟的一個特殊腔室,當心臟舒張時,血液通過側孔重新進入心臟,完成循環。
蜘蛛的血液,也就是血淋巴,是一種無色或淡藍色的液體。它的主要功能是運輸氧氣、營養物質和代謝廢物,但與脊椎動物不同,蜘蛛的血液不參與氧氣運輸。氧氣主要通過氣管系統直接輸送到組織。血淋巴中含有血細胞,這些細胞主要負責免疫功能,而不是氧氣運輸。
蜘蛛的循環系統展現出令人驚嘆的適應性。例如,某些生活在乾旱環境中的蜘蛛,如沙漠中的一些物種,能夠通過調節體液平衡來應對缺水環境。它們的血液中含有特殊的化合物,可以降低水分蒸發,幫助蜘蛛在高溫乾燥的環境中生存。
另一個有趣的適應是某些蜘蛛物種能夠利用循環系統來調節體溫。例如,某些日行性的跳蛛(Salticidae)在陽光下活動時,會增加心臟跳動頻率和血液流動速度,將熱量從曝露在陽光下的部位輸送到身體其他部分,從而調節體溫。
蜘蛛的循環系統還與其他生理系統密切相關。例如,在蛻皮過程中,循環系統起著關鍵作用。當蜘蛛準備蛻皮時,會增加體內液體壓力,這有助於舊外骨骼的脫落。此外,循環系統還參與蛛絲的生產過程,將必要的蛋白質和其他物質輸送到絲腺。
某些蜘蛛物種還表現出特殊的循環適應。例如,水蜘蛛(Argyroneta aquatica)能夠在水下生活,它們的循環系統經過特殊改造,能夠在水下環境中有效運作。這種蜘蛛能夠在水下建造充滿空氣的"鐘罩",並利用特殊的毛髮結構將氧氣運送到鐘罩中,從而維持水下呼吸。
研究蜘蛛的循環系統不僅有助於我們理解這些生物的生理機制,還為生物醫學領域提供了新的思路。例如,蜘蛛如何在開放式循環系統中有效控制體液平衡的研究,可能為開發新的水分調節藥物提供靈感。同時,蜘蛛在極端環境下的循環適應也為研究人類在特殊環境中的生理反應提供了重要參考。
5.3 蜘蛛毒液的產生與功能
蜘蛛毒液是這些八腳生物最為人知的特徵之一,它不僅在捕獵和防禦中扮演關鍵角色,還在醫學研究領域展現了巨大潛力。蜘蛛毒液的產生是一個複雜的生理過程,涉及專門的腺體和精密的輸送系統。
蜘蛛的毒腺通常位於頭胸部,由一對修飾過的外涎腺演化而來。這些腺體由特化的上皮細胞組成,能夠合成和分泌各種毒素。毒液的組成可能因蜘蛛種類而異,但通常包含多種生物活性物質,如多肽、蛋白質、生物胺和其他有機化合物。
毒液從毒腺產生後,通過導管輸送到螯肢(chelicerae)的尖端。螯肢是蜘蛛用於注射毒液的器官,類似於針筒的功能。當蜘蛛捕獵或防禦時,它們會用螯肢刺穿獵物或威脅者的表皮,同時釋放毒液。
蜘蛛毒液的主要功能是麻痺和消化獵物。許多毒素能夠干擾神經系統,導致獵物迅速癱瘓。這不僅有助於蜘蛛控制可能體型更大或更具攻擊性的獵物,還能防止獵物逃脫或反擊。此外,一些毒液成分具有消化作用,能夠開始分解獵物的組織,使蜘蛛更容易吸收營養。
值得注意的是,不同蜘蛛種類的毒液組成和效力各不相同。例如,澳洲的漏斗網蛛(funnel-web spider)以其對靈長類高度毒性的毒液聞名,而許多家庭常見的蜘蛛毒性則相對較弱。這種差異反映了蜘蛛在進化過程中對不同生態位的適應。
蜘蛛毒液在防禦中也發揮重要作用。雖然大多數蜘蛛傾向於逃避而非confrontation,但當面臨威脅時,毒液可以作為有效的威懾工具。某些蜘蛛種類,如黑寡婦(black widow),其毒液對脊椎動物也有強烈效果,這可能是為了應對更大型的捕食者。
近年來,蜘蛛毒液在醫學和農業領域引起了廣泛關注。研究人員發現,某些毒液成分具有潛在的藥用價值。例如,來自智利玫瑰蜘蛛(Chilean rose tarantula)的一種肽,顯示出可能用於治療慢性疼痛的前景。另一些毒素成分則在開發新型殺蟲劑方面展現了應用潛力,這可能為農業害蟲控制提供更環保的選擇。
蜘蛛毒液的產生是一個精確調控的過程。許多蜘蛛能夠根據不同情況調整毒液的組成和用量。例如,在面對大型獵物或感知威脅時,蜘蛛可能會分泌更多或更濃縮的毒液。相反,對於小型獵物,蜘蛛可能會節省毒液使用,因為產生毒液需要消耗大量能量。
值得一提的是,並非所有蜘蛛都依賴毒液捕獵。一些蜘蛛,如捕鳥蛛(bird-eating spider),主要依靠強大的螯肢和體型優勢來制服獵物。這反映了蜘蛛在捕獵策略上的多樣性適應。
蜘蛛毒液的研究不僅為我們提供了理解這些生物的獨特適應機制的窗口,還為生物醫學領域開闢了新的研究方向。隨著分析技術的進步,科學家們正在解析越來越多蜘蛛毒液的精細結構和功能,這可能導致新藥物的發現和開發。然而,重要的是要認識到,大多數蜘蛛對人類是無害的,它們的毒液主要用於生存,而非主動攻擊大型動物。
5.4 蜘蛛的體溫調節
蜘蛛作為變溫動物,其體溫調節機制是一個複雜而有趣的生理過程,直接影響著它們的生存和行為。儘管蜘蛛不像哺乳動物那樣能夠維持恆定的體溫,但它們已經演化出多種策略來應對環境溫度的變化,確保自身在不同氣候條件下都能正常運作。
蜘蛛的體溫主要受環境溫度影響,但它們並非完全被動地接受外界溫度的變化。相反,蜘蛛通過一系列行為和生理機制來調節體溫,以適應不同的溫度環境。這種調節能力對蜘蛛的生存至關重要,因為適當的體溫對它們的新陳代謝、活動能力和繁殖成功都有重大影響。
其中一個重要的體溫調節策略是微環境選擇。蜘蛛能夠感知周圍環境的溫度變化,並選擇最適合的棲息地。例如,在寒冷的季節,許多蜘蛛會尋找避風的角落或地下空間來過冬,這些地方溫度相對穩定,可以保護蜘蛛免受極端低溫的影響。相反,在炎熱的夏季,蜘蛛可能會選擇陰涼的樹蔭或岩石下方作為棲息地,以避免過高的溫度。
行為調節是蜘蛛體溫控制的另一個關鍵方面。當環境溫度過低時,一些蜘蛛會通過曬太陽來提高體溫。它們會將身體攤平,最大限度地暴露在陽光下以吸收熱量。有趣的是,一些沙漠蜘蛛甚至發展出了特殊的「蹺腳」行為,即在炎熱的沙地上抬起身體,減少與高溫地面的接觸,同時也能讓涼爽的空氣在腹部下方流通,起到降溫作用。
蜘蛛的體型和顏色也在體溫調節中扮演重要角色。較大的蜘蛛由於體表面積與體積比較小,保溫能力較強,而小型蜘蛛則更容易受環境溫度影響。深色的蜘蛛通常能更有效地吸收熱量,這在寒冷環境中是一種優勢。相反,淺色的蜘蛛則更容易反射熱量,有助於在高溫環境中保持涼爽。
一些蜘蛛種類還發展出了特殊的生理機制來應對極端溫度。例如,某些生活在寒冷地區的蜘蛛能夠在體內產生類似「防凍劑」的物質,降低體液的凝固點,防止在低溫環境中結冰。這種適應使它們能夠在零度以下的溫度中存活。另一方面,一些沙漠蜘蛛則進化出了特殊的水分保持機制,能夠在極度乾燥和炎熱的環境中維持體內水分平衡。
蜘蛛網的構建也與體溫調節有關。某些蜘蛛會根據環境溫度調整網的位置和結構。例如,在寒冷季節,一些蜘蛛會將網建在更靠近地面或植被茂密的地方,這些區域通常溫度較高且較為穩定。而在炎熱季節,它們可能會選擇更開闊、通風良好的地方來織網,以避免過熱。
值得注意的是,蜘蛛的體溫調節能力也影響著它們的活動模式。許多蜘蛛是夜行性的,這可能部分是為了避免白天的高溫。在溫帶地區,蜘蛛的活動通常在春秋季節最為活躍,因為這時的溫度最接近它們的最適溫度範圍。
蜘蛛的繁殖行為也與體溫調節密切相關。雌蜘蛛在孵化卵囊時,會選擇溫度適宜的位置,並通過調整身體姿勢來控制卵囊接收的熱量。某些蜘蛛甚至會在卵囊周圍織特殊的絲網結構,以調節溫度和濕度,為後代提供最佳的發育環境。
研究發現,氣候變化可能對蜘蛛的體溫調節能力產生重大影響。隨著全球氣溫升高,一些蜘蛛種類可能面臨適應新溫度環境的挑戰。這可能導致某些蜘蛛種類的分布範圍發生變化,甚至影響整個生態系統的平衡。
蜘蛛的體溫調節機制展示了自然界適應性進化的奇妙之處。通過結合行為策略、生理適應和微環境選擇,蜘蛛能夠在各種溫度條件下生存和繁衍。這種適應能力不僅確保了蜘蛛在地球上廣泛分布,也使它們成為研究氣候變化對生物影響的重要指標物種。隨著科技的進步,科學家們正在使用更精密的工具和方法來研究蜘蛛的體溫調節機制,這不僅有助於我們更好地理解這些神奇生物,也為生物仿生學和環境適應性研究提供了寶貴的見解。
6. 蛛網構築藝術
蛛網是蜘蛛建築藝術的巔峰之作,展現了自然界中的精巧設計和工程學奇蹟。蜘蛛利用其特殊的絲腺分泌蛛絲,這種材料兼具強度和彈性,是已知自然界中最堅韌的纖維之一。蜘蛛通過複雜的織網行為,將這些絲線編織成各種形狀和結構的網,每種網都有其獨特的功能和美學價值。
常見的蛛網類型包括輻射狀圓網、片狀網、漏斗網和不規則網等。其中,輻射狀圓網最為人熟知,如金園蛛(Nephila)所織的大型圓網就是典型代表。這種網結構優雅對稱,由放射狀的支撐絲和螺旋狀的黏絲組成,能有效捕捉飛行昆蟲。
蛛網不僅是捕獵工具,還是蜘蛛的家園。某些蜘蛛物種會在網中心或邊緣構建特殊的藏身處,用於躲避天敵和不利天氣。例如,園蛛(Argiope)常在網中心織一條之字形的裝飾帶,既可以穩定網結構,又能隱藏自身。
蛛網的建築藝術啟發了人類在多個領域的創新。建築師借鑒蛛網的結構原理設計輕質高強的建築,如北京國家體育場「鳥巢」就融入了蛛網的設計理念。此外,蛛絲的特性也促進了新型材料的研發,科學家正在努力研製人造蛛絲,期望應用於醫療、航空等領域。
6.1 蛛網的類型
蛛網是蜘蛛族群中最為獨特和引人注目的特徵之一,展現了蜘蛛在長期進化過程中所發展出的高度專業化和多樣化的捕獵策略。不同類型的蛛網反映了蜘蛛種類的多樣性,以及它們對各種生態環境的適應。每種蛛網都是其建造者的獨特簽名,揭示了蜘蛛的行為、生態位置和進化歷史。
最為人熟知的蛛網類型是輻射狀圓網。這種網由許多放射狀的支撐絲和螺旋狀的黏絲組成,形成一個圓形或橢圓形的結構。園蛛科(Araneidae)的成員,如十字蛛(Araneus)和金園蛛(Nephila),是這種網型的主要建造者。輻射狀圓網通常垂直懸掛,能有效捕捉飛行昆蟲。這種網的設計巧妙地利用了物理原理,當昆蟲撞擊網面時,網絲的彈性能夠吸收衝擊力,防止獵物反彈逃脫。同時,網面的振動能迅速傳遞到蜘蛛所在的位置,使其能快速反應前來捕獲獵物。
片狀網是另一種常見的蛛網類型,通常由錢蛛科(Linyphiidae)的成員所織。這種網呈水平或略微傾斜的片狀結構,由密集的不規則絲線組成。片狀網通常建在低矮植被間,適合捕捉從上方落下或從下方跳躍的小型昆蟲。有趣的是,一些錢蛛會在網的上方添加一些垂直的絲線,形成障礙物,使飛行昆蟲撞上後掉落到下方的片狀網中。
漏斗網是由漏斗蛛科(Agelenidae)蜘蛛所織,其特點是一個平坦的網面連接著一個漏斗狀的藏身處。這種網通常建在地面或低矮植被上,蜘蛛隱藏在漏斗深處,等待獵物踩踏網面時出來捕獲。漏斗網的結構不僅有利於捕獵,還為蜘蛛提供了良好的庇護所,保護其免受天敵和惡劣天氣的侵害。
不規則網,也稱為蓬網或迷宮網,是由許多看似雜亂無章的絲線組成的三維結構。這種網型常見於球蛛科(Theridiidae)的成員,如黑寡婦蜘蛛(Latrodectus)。不規則網雖然看起來雜亂,但實際上結構複雜,能有效捕獲各種類型的昆蟲。當獵物觸碰到網的任何部分,都可能被纏住或掉落到網的下部,在掙扎中越陷越深。
管狀網是另一種獨特的蛛網類型,通常由管居蛛科(Atypidae)的成員所織。這種網呈管狀,一端封閉,另一端開口,蜘蛛棲息在管內。當獵物經過管的外表面時,蜘蛛會感知到振動,迅速咬破管壁捕獲獵物。這種網型不僅是捕獵工具,也是蜘蛛的家園,提供了良好的保護和隱蔽。
蓑網是一種相對罕見但極具特色的蛛網類型,由三角蛛科(Uloboridae)的某些成員所織。這種網看起來像一個小型的吊床或蓑衣,通常懸掛在樹枝或其他植物結構上。蓑網的特殊形狀使它能夠捕獲特定類型的飛行昆蟲,展現了蜘蛛對特定生態位的適應。
值得一提的是,並非所有蜘蛛都織網捕獵。有些蜘蛛,如狼蛛科(Lycosidae)和跳蛛科(Salticidae)的成員,主要依靠敏捷的行動能力和鋒利的視力來追捕獵物。這些蜘蛛可能只使用絲線來製作簡單的藏身處或用於繁殖目的,而不是捕獵。
某些蜘蛛物種還發展出了特殊的網型變體或獨特的使用方式。例如,有些園蛛會在網中添加之字形的裝飾帶,可能用於穩定網結構或吸引獵物。另一個有趣的例子是幽靈蛛屬(Deinopis)的成員,它們織造一種小型的網,握在前肢上,當獵物接近時迅速將網拋出捕獲。
蛛網的多樣性不僅體現在結構上,還反映在材料使用上。不同的蜘蛛物種能夠產生不同性質的絲,有些絲具有極強的黏性,而有些則更注重彈性和強度。這種材料的多樣性使得蜘蛛能夠根據自身需求和環境條件來調整網的性能。
蛛網類型的研究不僅對理解蜘蛛的生態和行為至關重要,還為生物材料科學和仿生工程提供了寶貴的靈感。科學家們正在深入研究蛛網的結構和材料特性,希望能夠開發出新型的高性能材料和結構設計。
6.2 蛛網的功能
蛛網是蜘蛛生存策略中的核心元素,其功能遠遠超出了人們通常認知的簡單捕獵工具。事實上,蛛網在蜘蛛的生活中扮演著多重角色,展現了自然界中精巧的多功能設計。
首要而言,蛛網最為人知的功能是捕獵。蛛網作為一個高效的捕獵裝置,能夠大大擴展蜘蛛的捕獵範圍。以常見的輻射狀圓網為例,它能夠攔截大範圍內的飛行昆蟲。網絲的黏性和彈性使得獵物一旦接觸就難以脫身,而網絲的震動又能迅速通知蜘蛛獵物的位置。一些蜘蛛物種,如金園蛛(Nephila),其網可以達到一米以上的直徑,能夠捕獲包括蝴蝶和蜻蜓在內的較大型昆蟲。不同類型的蛛網適應於捕獲不同的獵物,例如,片狀網更適合捕捉跳躍的小型昆蟲,而不規則網則能夠纏住各種大小和類型的獵物。
除了捕獵,蛛網還具有重要的防禦功能。許多蜘蛛將蛛網作為隱蔽和保護自身的屏障。例如,漏斗蛛(Agelenidae)織造的漏斗網不僅是捕獵裝置,漏斗狀的一端還為蜘蛛提供了一個安全的藏身之處,使其能夠躲避天敵的襲擊。一些蜘蛛物種甚至會在網上添加特殊的結構,如園蛛(Argiope)常在網中心織造醒目的之字形裝飾帶,這不僅可能用於吸引獵物,還可能起到混淆天敵視線的作用,保護蜘蛛免受攻擊。
蛛網在蜘蛛的溝通系統中也扮演著關鍵角色。對許多蜘蛛來說,蛛網是一個複雜的信息傳遞平台。雄性蜘蛛在求偶時常常通過撥動雌性的網絲來傳遞信號,這種振動模式可能包含了種族識別、求偶意圖等重要信息。一些蜘蛛物種還會利用網絲來傳遞警告信號,當感受到威脅時,它們會快速抖動整個網體,警告同類或嚇退潛在的捕食者。
在繁殖過程中,蛛網同樣發揮著重要作用。許多蜘蛛物種利用絲線來製作保護卵囊的結構。例如,球蛛科(Theridiidae)的成員常常將卵囊懸掛在不規則網的上部,既能保護卵囊免受天敵侵害,又能調節溫度和濕度,為後代提供理想的孵化環境。一些蜘蛛甚至會在網上特意留出空間作為幼蛛的生長區域,幫助幼蛛在初期獲得充足的食物和保護。
蛛網還在蜘蛛的遷移和擴散中起著關鍵作用。許多幼蛛和小型蜘蛛會釋放長絲進行「飛行」,這種行為被稱為「飄絲」。通過這種方式,蜘蛛可以藉助風力遷移到新的棲息地,有效擴大種群的分布範圍。這種擴散策略對於維持種群基因多樣性和適應新環境至關重要。
在環境感知方面,蛛網作為蜘蛛的「延伸感官」發揮著重要作用。蜘蛛通過網絲的震動可以感知周圍環境的變化,包括獵物的出現、天敵的接近,甚至是天氣的變化。一些研究表明,蜘蛛可能通過網絲的張力變化來預測即將到來的降雨或強風,從而及時加固網結構或尋找庇護所。
蛛網在蜘蛛的能量管理中也扮演著重要角色。織網是一項耗能的活動,因此許多蜘蛛會「回收利用」舊的網絲。當網絲損壞或效用降低時,蜘蛛會將其吃掉,重新吸收其中的蛋白質,用於新網絲的生產。這種循環利用的策略使得蜘蛛能夠在資源有限的情況下維持高效的捕獵活動。
某些蜘蛛物種還將蛛網用於特殊的功能。例如,某些水生蜘蛛會利用網絲在水下構建充滿空氣的「潛水鐘」,作為水下的居所和獵場。這種獨特的適應使得這些蜘蛛能夠在水生環境中生存和繁衍。
蛛網的多功能性不僅體現在蜘蛛的生存策略中,還為人類提供了寶貴的研究靈感。生物材料科學家正在研究蛛絲的獨特性質,希望開發出新型的高強度、高彈性材料。建築學家和工程師則從蛛網的結構設計中汲取靈感,應用於建築和橋樑等大型結構的設計中。
6.3 織網行為
蜘蛛的織網行為是一個複雜而精密的過程,展現了這些小生物驚人的工程學天賦和本能行為的精確性。每個蜘蛛物種都有其獨特的織網方式,但大多數織網行為都遵循一些基本的原則和步驟。
以最常見的輻射狀圓網為例,織網過程通常始於架設框架線。蜘蛛會先選擇一個合適的位置,這個選擇涉及多個因素,包括周圍的植被結構、氣流patterns、潛在獵物的活動範圍等。一旦選定位置,蜘蛛會釋放一根絲線,讓風將其吹向預定的錨點。當絲線附著後,蜘蛛會沿著這根絲線移動,同時分泌第二根絲線以加強結構。這個過程會重複多次,直到形成一個基本的多邊形框架。
接下來,蜘蛛會在框架內部織造輻射狀的支撐絲。這些絲線從網的中心向外延伸,連接到框架上。支撐絲的數量和分佈會根據蜘蛛物種和網的大小而有所不同。例如,園蛛科(Araneidae)的成員通常會織造20到30根支撐絲,而金園蛛(Nephila)的大型網絡可能包含更多支撐絲。
在完成支撐絲的構建後,蜘蛛會開始織造捕獲絲,也就是螺旋狀的黏性絲線。這個過程通常從外圍開始,蜘蛛沿著支撐絲移動,逐漸向中心螺旋前進。有趣的是,許多蜘蛛在織造黏性捕獲絲之前,會先織一個臨時的非黏性螺旋,作為腳架輔助構建。當黏性螺旋完成後,這個臨時螺旋通常會被拆除。
整個織網過程展現了蜘蛛驚人的空間感和幾何學直覺。研究表明,蜘蛛能夠根據環境條件和自身需求調整網的大小和形狀。例如,在風力較大的環境中,蜘蛛可能會減少網的大小或增加支撐絲的數量以增強穩定性。
不同物種的蜘蛛在織網行為上也有顯著差異。例如,某些園蛛物種會在網中添加醒目的之字形或圓形裝飾帶,這種行為的確切功能仍有爭議,可能用於穩定網結構、吸引獵物或混淆天敵視線。另一個有趣的例子是三角蛛科(Uloboridae)的成員,它們會織造特殊的蓑網,這種網看起來像一個小型吊床,需要非常精確的織造技巧。
織網行為還受到多種外部因素的影響。光照條件是一個重要因素,許多蜘蛛物種傾向於在黎明或黃昏時分織網,這可能是為了避免被天敵發現或適應獵物的活動規律。氣象條件也會影響織網行為,例如,一些蜘蛛在感知到即將有強風或降雨時,會選擇延遲織網或構建更加堅固的網結構。
蜘蛛的年齡和生理狀態也會影響其織網行為。年輕的蜘蛛通常織造較小的網,隨著年齡增長和經驗積累,網的大小和複雜度會逐漸增加。此外,雌性蜘蛛在懷卵期間可能會改變織網習慣,有些物種會減少織網頻率或改變網的結構以適應新的生理需求。
值得注意的是,蜘蛛的織網行為並非完全固定不變。研究發現,蜘蛛具有一定的學習能力和適應性。例如,當蜘蛛反複遇到某種特定類型的獵物時,它們可能會微調網的結構以提高捕獲效率。同樣,如果某個位置長期缺乏獵物,蜘蛛可能會選擇更換織網地點。
織網行為還涉及複雜的能量平衡。織網是一項耗能的活動,因此蜘蛛需要權衡織網的成本和潛在收益。一些蜘蛛物種會定期更換整個網絡,而另一些則傾向於修補和維護現有的網。這種選擇可能取決於環境因素、獵物豐富程度以及蜘蛛自身的能量儲備。
某些蜘蛛物種還展現出協作織網的行為。例如,社會性蜘蛛科(Anelosimus)的成員會共同織造大型的集群網絡,這種行為不僅提高了捕獵效率,還為整個族群提供了保護。
蜘蛛的織網行為一直是生物學家和工程師關注的焦點。通過研究蜘蛛如何在複雜的三維空間中構建精密的網絡結構,科學家們希望能夠開發出新的建築技術和材料設計方法。同時,蜘蛛織網過程中展現的精確性和適應性也為人工智能和機器人技術的發展提供了寶貴的啟示。
6.4 蛛絲的特性與用途
蛛絲是自然界中最令人驚嘆的材料之一,其獨特的物理和化學特性使其成為科學研究和技術創新的重要對象。蛛絲的主要成分是蛋白質,但其結構和性質遠非普通蛋白質可比。蜘蛛能夠產生多種不同類型的絲,每種絲都有其特定的功能和特性。
蛛絲最為人知的特性是其驚人的強度和韌性。就重量而言,蛛絲比鋼鐵更強韌,同時又具有極高的彈性。這種獨特的組合使得蛛絲能夠吸收巨大的衝擊力而不斷裂。例如,金園蛛(Nephila)的拖曳絲強度可達到高碳鋼的五倍,同時其延展性可達到尼龍的兩倍。這種特性使得蛛網能夠有效捕獲高速飛行的昆蟲而不被撕裂。
除了強度和韌性,蛛絲還具有優異的熱穩定性和化學穩定性。在寬廣的溫度範圍內,蛛絲能夠保持其物理特性不變。同時,蛛絲對多種化學物質具有抗性,這使得它在自然環境中能夠長期保持功能。這種穩定性源於蛛絲蛋白質的獨特分子結構,其中包含了大量的β-折叠片層結構,這種結構賦予了蛛絲高度的穩定性和強度。
蛛絲還具有出色的生物相容性和可降解性。與許多合成材料不同,蛛絲在生物體內不會引起明顯的免疫反應,同時又能夠被自然分解。這種特性使得蛛絲在醫療領域具有廣泛的應用前景,例如用於製作手術縫合線或組織工程支架。
另一個值得注意的特性是蛛絲的黏性。某些類型的蛛絲,如圓網蜘蛛的捕獲絲,具有極強的黏性,能夠有效粘住獵物。這種黏性來源於絲上的特殊黏液滴,這些黏液滴不僅能夠粘住獵物,還能夠在濕度變化時保持黏性。這種特性啟發了新型黏合劑的研發。
蛛絲的多功能性體現在蜘蛛使用不同類型絲的方式上。例如,抓絲用於構建網的主體結構,拖曳絲用於蜘蛛在空中移動,而包裹絲則用於包裹獵物。每種絲都有其特定的物理和化學特性,適應不同的功能需求。
在自然界中,蛛絲主要用於蜘蛛的捕獵、移動和繁殖。然而,人類對蛛絲的潛在應用產生了濃厚的興趣。在醫療領域,蛛絲被視為製作手術縫合線的理想材料,因為它既強韌又能被人體吸收。此外,蛛絲的生物相容性使其成為組織工程和藥物遞送系統的潛在候選材料。
在工業領域,科學家們正在研究如何利用蛛絲的特性開發新型材料。例如,仿生蛛絲可能用於製作輕質高強的防彈衣、降落傘或飛機零件。蛛絲的彈性特性也使其成為運動器材和彈性纖維的潛在原料。
在環境保護方面,蛛絲作為一種可生物降解的天然材料,可能為解決塑料污染問題提供新的思路。研究人員正在探索使用仿生蛛絲替代某些不可降解的合成材料。
然而,大規模生產蛛絲面臨著巨大挑戰。與蠶絲不同,蜘蛛難以大規模飼養,因為它們往往具有領地性和食同類行為。因此,科學家們轉向了基因工程和化學合成的方法。通過在細菌、酵母甚至山羊中引入蜘蛛絲基因,研究人員希望能夠大量生產具有類似特性的蛋白質。
蛛絲的研究不僅局限於材料科學,還涉及生物學、化學和物理學等多個領域。了解蛛絲形成的分子機制可能為蛋白質折疊和自組裝研究提供新的見解。同時,蛛絲的力學特性研究也為材料科學和工程領域提供了寶貴的啟示。
值得一提的是,不同蜘蛛物種產生的絲可能具有不同的特性。例如,達爾文樹皮蛛(Caerostris darwini)產生的絲被認為是自然界中最強韌的生物材料之一,其強度幾乎是凱夫拉纖維的十倍。這種多樣性為研究人員提供了豐富的研究素材,有助於開發具有不同特性的新型材料。
6.5 台灣常見的蛛網類型
台灣有多種蜘蛛物種,這些蜘蛛織造出各式各樣的蛛網。這些蛛網不僅反映了蜘蛛的多樣性,也展現了蜘蛛對台灣獨特環境的適應。
在台灣最常見的蛛網類型之一是輻射狀圓網。這種網型主要由園蛛科(Araneidae)的成員所織造,如臺灣中型金蛛(Nephila pilipes)和臺灣銀腹蛛(Argiope aemula)。這些蛛網通常懸掛在開闊的空間中,如樹枝間或建築物的角落。輻射狀圓網的特點是由多根輻射狀的支撐絲和螺旋狀的捕獲絲組成,形成一個圓形或橢圓形的結構。這種網型非常適合捕捉飛行昆蟲,在台灣的森林、公園和郊區都能常見。臺灣中型金蛛的網可以達到一米以上的直徑,是台灣最大的蛛網之一,常常引起路人的注意。
片狀網是另一種在台灣常見的蛛網類型,主要由錢蛛科(Linyphiidae)的成員所織造。這種網通常呈水平或略微傾斜的片狀結構,由密集的不規則絲線組成。在台灣的低矮植被中,如草叢或灌木叢中,經常可以看到這種網型。片狀網特別適合捕捉從植物上跳躍或掉落的小型昆蟲。台灣的高山地區,如玉山或合歡山的草原地帶,常常可以看到大量的片狀網分布其中。
不規則網,也稱為蓬網或迷宮網,在台灣也相當普遍。這種網型主要由球蛛科(Theridiidae)的成員所織造,如黑寡婦蛛屬(Latrodectus)的物種。不規則網看似雜亂無章,實際上是一個複雜的三維結構,能有效捕獲各種類型的昆蟲。在台灣的建築物角落、樹幹縫隙或堆積的雜物中,常常可以發現這種網型。台灣的亞熱帶氣候為這類蜘蛛提供了理想的生存環境,使得不規則網在都市和鄉村地區都相當常見。
漏斗網是台灣另一種常見的蛛網類型,主要由漏斗蛛科(Agelenidae)的成員所織造。這種網的特點是一個平坦的網面連接著一個漏斗狀的藏身處。在台灣的地面植被、低矮灌木叢中,甚至是建築物的角落,都可能看到漏斗網的蹤跡。這種網型不僅是有效的捕獵工具,還為蜘蛛提供了良好的庇護所。在台灣的濕地或森林邊緣地帶,漏斗網尤為常見。
臺灣的亞熱帶環境也適合一些特殊的蛛網類型生存。例如,由鳥蛛科(Theraphosidae)的某些成員所織造的管狀網。雖然鳥蛛並不以織網聞名,但某些物種會在地面或樹幹上織造簡單的管狀絲巢。這種網型在台灣的低海拔森林地區可以觀察到,雖然相對罕見,但卻是台灣蛛網多樣性的重要組成部分。
在台灣的水域附近,如溪流、池塘或水稻田邊,可以看到一種獨特的網型——鐘形網。這種網主要由水蛛科(Cybaeidae)的成員所織造。水蛛會在水下織造一個鐘形的網,並填充空氣,形成一個水下的空氣室。這種適應使得這類蜘蛛能夠在水生環境中生存和捕獵。
台灣的海岸地區和離島,如墾丁或綠島,還可以觀察到一種特殊的蛛網——蓑網。這種網型主要由三角蛛科(Uloboridae)的成員所織造,形狀類似一個小型的吊床或蓑衣。蓑網通常懸掛在灌木或岩石上,特別適合捕捉飛行昆蟲。
值得一提的是,台灣的城市環境也為某些蜘蛛物種提供了獨特的織網場所。例如,在台北、高雄等大城市的高樓大廈之間,有些蜘蛛已經適應了在金屬和玻璃結構上織網。這種適應性展示了蜘蛛對都市化環境的驚人適應能力。
台灣的季節變化雖然不如溫帶地區明顯,但仍然會影響某些蛛網的分布和特徵。例如,在乾季時,某些蜘蛛可能會選擇在更潮濕的微環境中織網,而在雨季,一些物種可能會增加網的大小以捕捉更多的飛行昆蟲。
此外,台灣特有的地理環境,如高山地區和濱海地帶,也造就了一些獨特的蛛網適應。高山地區的蛛網通常需要更強的結構以抵抗強風,而濱海地區的蛛網則可能具有更好的耐鹽性。
總的來說,台灣豐富多樣的蛛網類型不僅反映了該島嶼的生物多樣性,也展示了蜘蛛對不同生態環境的適應能力。這些蛛網不僅是生態系統中重要的一環,也為研究人員提供了豐富的研究素材,有助於我們更好地理解蜘蛛的行為生態和進化適應。
7. 蜘蛛的行為生態
蜘蛛的行為生態是一個豐富多樣的研究領域,涵蓋了這些八腳動物在自然環境中的各種行為模式和生存策略。蜘蛛的行為生態主要包括覓食、防禦、繁殖和社會行為等方面。
在覓食方面,蜘蛛展現出多種獨特的策略。有些蜘蛛依賴織網捕獲獵物,如園蛛科(Araneidae)的成員;而其他如跳蛛科(Salticidae)的蜘蛛則採用主動獵捕的方式。蜘蛛的覓食行為常常涉及複雜的決策過程,包括選擇織網位置、調整網的結構以及對不同獵物採取不同的捕獲策略。
防禦行為是蜘蛛生存的另一個重要方面。許多蜘蛛物種 演化出了各種偽裝技巧,如擬態枯葉或鳥糞。一些蜘蛛還會在網上添加特殊的結構,如警戒線或裝飾帶,以混淆捕食者。此外,某些蜘蛛在受到威脅時會表現出特殊的行為,如假死或快速逃跑。
蜘蛛的繁殖行為也十分引人注目,尤其是求偶過程。雄性蜘蛛常常需要進行複雜的求偶舞蹈以避免被雌性當作獵物吃掉。某些物種的雄性會給雌性帶來「求偶禮物」,通常是包裹在絲中的獵物。
儘管大多數蜘蛛是獨居動物,但也有一些物種表現出社會行為。例如,某些蜘蛛物種會形成大型的集群網,共同捕獵和分享獵物。這種社會行為在資源豐富的環境中特別常見,如熱帶雨林地區。
7.1 蜘蛛的覓食策略
蜘蛛的覓食策略是蜘蛛行為生態學中最為引人入勝的方面之一,展現了這些生物在長期進化過程中發展出的多樣化和高度專業化的捕食技巧。蜘蛛的覓食策略大致可以分為兩大類:織網捕獵和主動獵捕,但在這兩大類別中又存在諸多變體和特殊適應。
織網捕獵是最為人熟知的蜘蛛覓食策略。採用這種策略的蜘蛛物種通常會在特定位置構建精巧的網狀結構,利用網的黏性和彈性來捕獲飛行或跳躍的獵物。園蛛科(Araneidae)的成員是這種策略的典型代表。例如,金園蛛(Nephila)會織造大型的輻射狀圓網,這種網不僅能夠捕獲小型昆蟲,甚至能夠捕獲體型較大的飛蛾和蝴蝶。
織網捕獵策略的一個關鍵要素是網站的選擇。蜘蛛需要權衡多個因素,如周圍的植被結構、氣流patterns、潛在獵物的活動範圍等。研究發現,經驗豐富的蜘蛛通常能夠選擇獵物更為豐富的位置來織網。此外,一些蜘蛛物種還會根據環境條件調整網的結構。例如,在風力較大的環境中,蜘蛛可能會減少網的大小或增加支撐絲的數量以提高穩定性。
某些採用織網策略的蜘蛛還發展出了特殊的捕獵技巧。例如,黏網蛛科(Theridiidae)的成員會在網的下方懸掛黏性垂絲。當獵物接觸這些垂絲時,蜘蛛會迅速將獵物拉起,使其遠離地面而無法逃脫。另一個有趣的例子是竹蛛屬(Pholcus)的蜘蛛,它們會快速旋轉身體,使網產生振動,纏繞並制服獵物。
與織網捕獵相對的是主動獵捕策略。採用這種策略的蜘蛛不依賴蛛網,而是主動搜尋和追捕獵物。跳蛛科(Salticidae)是這種策略的典型代表。這些蜘蛛擁有極為發達的視覺系統,能夠精確定位獵物,然後通過快速跳躍來捕獲它們。跳蛛的捕獵行為展現了驚人的計算能力和空間感,它們能夠準確估算到獵物的距離和位置,並在跳躍時作出必要的調整。
螃蟹蛛科(Thomisidae)的成員則採用了一種獨特的伏擊策略。這些蜘蛛通常棲息在花朵或樹葉上,靜待獵物接近。它們的體色常常與周圍環境相似,能夠有效地偽裝自己。當獵物靠近時,螃蟹蛛會突然伸出前肢抓住獵物。這種策略特別適合捕捉蝴蝶、蜜蜂等授粉昆蟲。
狼蛛科(Lycosidae)的蜘蛛則展現了另一種主動獵捕策略。這些蜘蛛通常在地面上快速移動,追逐並捕獲各種地棲昆蟲。狼蛛的視力雖然不如跳蛛,但它們擁有敏銳的震動感應能力,能夠通過地面振動來定位獵物。
某些蜘蛛物種還發展出了特殊的覓食策略。例如,漁蛛屬(Dolomedes)的成員能夠在水面上行走,甚至潛入水中捕獲小魚或水生昆蟲。這種獨特的適應使得它們能夠利用水生環境這一特殊的生態位。
值得注意的是,許多蜘蛛物種的覓食策略會隨年齡和性別而變化。例如,某些網織蛛的幼體可能主要依賴主動獵捕,而成年個體則轉向織網捕獵。同樣,在某些物種中,雄性和雌性可能採用不同的覓食策略,這通常與它們的體型差異和能量需求有關。
蜘蛛的覓食策略還涉及複雜的能量平衡考量。織網捕獵雖然能夠捕獲大量獵物,但織網本身是一項耗能的活動。因此,蜘蛛需要權衡織網的成本和潛在收益。某些蜘蛛物種會根據獵物的豐富程度來調整織網頻率,在食物稀缺時減少織網以節省能量。
此外,一些蜘蛛物種還表現出食物偏好和選擇性。研究發現,某些蜘蛛能夠區分不同類型的獵物,並根據獵物的營養價值和捕獲難度來做出選擇。這種選擇性捕食行為有助於蜘蛛最大化其能量攝入。
蜘蛛的覓食策略還受到環境因素的影響。例如,在城市環境中,某些蜘蛛物種已經適應了利用人造光源來增加捕獲夜行性昆蟲的機會。這種行為適應展示了蜘蛛對人為環境變化的回應能力。
7.2 蜘蛛如何避免被捕食
蜘蛛作為生態系統中的重要捕食者,同時也面臨著來自其他動物的捕食壓力。為了應對這種生存挑戰,蜘蛛在長期進化過程中發展出了一系列複雜而有效的避免被捕食的策略。這些策略涵蓋了行為、形態和生理等多個方面,展現了蜘蛛在生存競爭中的適應能力。
偽裝是蜘蛛最常見且最有效的防禦策略之一。許多蜘蛛物種能夠通過改變體色或形態來融入周圍環境,從而避免被天敵發現。例如,螃蟹蛛科(Thomisidae)的成員常常棲息在花朵上,它們的體色可以是白色、黃色或粉色,與花瓣顏色完美匹配。另一個典型例子是枯葉蛛(Poltys), 它們的外形和顏色酷似枯葉,當停留在樹枝上時幾乎無法被察覺。
除了靜態偽裝,一些蜘蛛還能夠進行動態偽裝。例如,某些園蛛物種在受到驚擾時會快速擺動身體,使自己的輪廓變得模糊,這種行為被稱為「搖擺行為」。這種動態偽裝不僅能夠混淆捕食者的視覺,還可能使捕食者誤以為蜘蛛是不可食用的物體。
警戒色彩是另一種有趣的防禦策略。某些蜘蛛物種,如南美洲的孔雀蛛(Maratus),擁有鮮艷的體色。這種顯眼的外表通常被解釋為對捕食者的警告信號,暗示蜘蛛可能有毒或味道不佳。雖然大多數蜘蛛實際上對大型捕食者並無危害,但這種虛張聲勢的策略往往能夠有效地嚇退潛在的威脅。
某些蜘蛛物種還發展出了特殊的行為來避免被捕食。例如,某些園蛛在白天會將自己縮成一團,隱藏在樹葉背面或樹皮縫隙中。到了夜晚,它們才會重新展開身體,開始活動和織網。這種晝伏夜出的行為可以有效減少被日間活動的鳥類等捕食者發現的風險。
跳蛛科(Salticidae)的成員則展現出了更為主動的防禦行為。這些蜘蛛擁有極為發達的視覺系統,能夠迅速察覺潛在的威脅。當感受到危險時,跳蛛可以快速跳躍逃離,其敏捷的身手常常能夠讓捕食者措手不及。
某些蜘蛛物種還利用自身的絲來增強防禦能力。例如,某些園蛛會在網上添加特殊的結構,如之字形的裝飾帶或螺旋狀的絲線。這些結構不僅可能用於穩定網體,還可能起到混淆捕食者視線的作用。另一個有趣的例子是某些熱帶蜘蛛物種,它們會在網周圍織造大量的不規則絲線,形成一個「絲網屏障」,這不僅能夠阻擋捕食者的接近,還可能使捕食者被纏住。
在面對直接威脅時,許多蜘蛛物種會採取「裝死」的策略。當受到驚擾時,蜘蛛會突然停止所有動作,將腳緊緊蜷縮在身體周圍。這種行為可能欺騙捕食者,使其誤以為蜘蛛已經死亡或不具有威脅。一旦危險過去,蜘蛛就會迅速「復活」並逃離。
某些蜘蛛物種還具有自斷肢體的能力。當被捕食者抓住腿部時,蜘蛛可以主動斷開該肢體,從而逃脫危險。雖然這種策略會導致暫時的行動不便,但對於保住性命來說是一個值得的權衡。更令人驚奇的是,許多蜘蛛物種能夠在後續的蛻皮過程中重新長出失去的肢體。
化學防禦也是某些蜘蛛物種採用的策略。例如,某些蜘蛛物種能夠分泌具有刺激性或難聞氣味的物質,這些物質可以嚇退潛在的捕食者。雖然大多數蜘蛛的毒液主要用於麻痺獵物,但對於某些大型蜘蛛物種來說,毒液也可能成為對抗捕食者的有力武器。
社會性行為在某些蜘蛛物種中也起到了重要的防禦作用。例如,某些社會性蜘蛛物種會共同織造大型的集群網,這不僅提高了捕獲獵物的效率,還為整個群體提供了更好的保護。當面臨威脅時,群體中的個體可以互相警戒,增加察覺危險的機會。
值得注意的是,蜘蛛的防禦策略常常是多方面的綜合運用,而不是單一的方法。一個蜘蛛物種可能同時具備偽裝能力、警戒行為和逃跑技巧。這種多元化的防禦策略使得蜘蛛能夠應對各種不同類型的捕食威脅,從而在複雜的生態系統中生存和繁衍。
7.3 蜘蛛的社會行為
蜘蛛通常被認為是獨居的生物,但實際上,某些蜘蛛物種展現出令人驚訝的社會行為。這種社會性在蜘蛛中雖然相對罕見,但卻提供了豐富的研究素材,有助於我們理解社會行為的演化過程和生態意義。
社會性蜘蛛的最顯著特徵是它們共同生活和協作捕獵的能力。這種行為在熱帶和亞熱帶地區的某些物種中尤為常見。例如,社會性蛛科(Anelosimus)的一些成員,如厄瓜多爾社會性蛛(Anelosimus eximius),能夠形成由數百甚至上千個個體組成的巨大群落。這些蜘蛛共同織造一個大型的網絡結構,可以覆蓋整棵樹或大片灌木。
在這種大型集群網中,社會性蜘蛛展現出高度的協作行為。當獵物落入網中時,多個蜘蛛會同時攻擊,共同將獵物制服。這種協作捕獵行為使得它們能夠捕獲比自身大得多的獵物,大大提高了捕食效率。捕獲的獵物會被帶回巢穴中心,由群體成員共同分享。這種食物分享行為是社會性蜘蛛一個重要的特徵,表現出了原始的資源分配機制。
社會性蜘蛛的另一個顯著特徵是共同育幼。在許多社會性蜘蛛群體中,照顧幼蛛不僅僅是母親的責任,其他成年個體也會參與其中。這種共同育幼行為大大提高了幼蛛的存活率。例如,在某些社會性蛛物種中,成年個體會輪流照看卵囊,確保它們處於適宜的溫度和濕度環境中。當幼蛛孵化後,群體中的成年個體會共同為它們提供食物,甚至進行餵食。
有趣的是,社會性蜘蛛群體中通常存在分工現象。雖然不像社會性昆蟲那樣有明確的階級制度,但不同個體可能會傾向於執行不同的任務。例如,某些個體可能更多地參與織網和修補網絡,而另一些則主要負責捕獵或育幼。這種非正式的分工有助於提高整個群體的效率。
社會性蜘蛛的群體還表現出集體防禦行為。當面臨威脅時,群體中的個體會協同行動以驅趕入侵者。例如,當寄生蜂試圖入侵巢穴時,多個蜘蛛會同時攻擊,形成一道有效的防線。這種集體防禦不僅保護了成年個體,更重要的是保護了脆弱的幼蛛和卵囊。
然而,社會性行為在蜘蛛中並非全有全無的現象,而是存在一個連續譜。在這個譜系的一端是高度社會性的物種,如前面提到的厄瓜多爾社會性蛛;而在另一端則是完全獨居的物種。在這兩個極端之間,存在著各種程度的社會性。
例如,某些蜘蛛物種展現出「亞社會性」行為。這些蜘蛛雖然不形成永久性的大型群落,但會在特定時期表現出一定程度的社會性。比如,某些園蛛物種的幼體在剛孵化後的一段時間內會聚集在一起,共同織網和捕獵。隨著年齡增長,它們逐漸分散,過渡到獨居生活。
另一種有趣的社會行為形式是「周期性社會性」。某些蜘蛛物種在特定季節或環境條件下會形成臨時的群體。例如,在食物特別豐富的地方,一些通常獨居的蜘蛛可能會聚集在一起,共同捕獵和分享食物。這種行為可能是對環境資源分布不均的一種適應。
值得注意的是,社會性行為的演化可能受到多種因素的影響。研究表明,資源可用性、捕食壓力、氣候條件等環境因素可能推動社會性的發展。同時,遺傳因素也在其中扮演重要角色。某些研究發現,社會性蜘蛛往往具有較高的近親繁殖傾向,這可能有助於維持群體內的合作行為。
社會性蜘蛛的研究不僅對理解蜘蛛行為生態學有重要意義,還為我們提供了研究社會行為演化的獨特視角。與社會性昆蟲不同,社會性蜘蛛沒有明顯的繁殖分工,這使得它們成為研究社會行為早期演化階段的理想模型。
此外,社會性蜘蛛的研究還可能對生態系統管理和生物防治領域產生影響。例如,了解社會性蜘蛛的群體動態和捕食行為,可能有助於開發更有效的生物防治策略,用於控制農業害蟲。
7.4 蜘蛛的通訊方式
蜘蛛的通訊方式是一個引人入勝的主題,涵蓋了多種感官和行為適應。儘管蜘蛛通常被視為獨居生物,但它們實際上擁有複雜的通訊系統,用於各種社交互動。
蜘蛛主要通過振動、化學和視覺信號進行通訊。振動通訊是最普遍的形式,特別是在織網蜘蛛中。蜘蛛利用網絲傳遞振動信號,這些信號可以傳達多種信息,如求偶意圖、警告或領地宣示。例如,雄性蜘蛛在求偶時會在雌性的網上產生特定的振動模式,以表明自己的存在和意圖。這些振動模式通常是物種特異性的,有助於避免種間混淆。
化學通訊在蜘蛛中也扮演著重要角色,尤其是在性信息素的傳遞上。雌性蜘蛛會釋放特定的化學物質來吸引雄性。這些信息素可以在空氣中傳播,或附著在蛛絲上。一些游走蜘蛛物種(種)還會使用化學標記來標識它們的領地或追蹤獵物。
視覺通訊在某些蜘蛛物種(種)中也很重要,特別是在跳蛛(Salticidae)家族中。雄性跳蛛often(常常)會進行複雜的求偶舞蹈,展示他們鮮艷的體色和特殊的肢體動作來吸引雌性。這種視覺display(展示)不僅用於求偶,也可用於種間識別和領地防衛。
觸覺通訊在蜘蛛的近距離互動中也起著重要作用。蜘蛛利用它們的腳和觸肢來感知和傳遞信息。例如,在交配過程中,雄性蜘蛛會使用特定的觸摸模式來安撫雌性,降低被捕食的風險。
一些蜘蛛物種(種)還能夠產生聲音進行通訊。例如,某些狼蛛物種(種)能夠通過摩擦身體部位產生可聽見的聲音,這可能用於驅趕捕食者或吸引配偶。
蜘蛛的通訊系統也適應了它們的生態位。例如,生活在密集植被中的蜘蛛可能更依賴化學和觸覺信號,而生活在開闊地帶的蜘蛛則可能更多地利用視覺信號。
有趣的是,一些蜘蛛物種(種)還能夠利用其他動物的通訊系統。例如,某些蟹蛛物種(種)能夠模仿昆蟲的信息素,藉此吸引獵物靠近。這種欺騙性通訊策略展示了蜘蛛通訊系統的複雜性和適應性。
在社會性蜘蛛中,通訊系統更為複雜。這些蜘蛛需要協調群體活動,如共同織網、獵食和育幼。它們可能使用一系列的振動和化學信號來維持社群凝聚力和組織集體行為。
蜘蛛的通訊方式也受到環境因素的影響。例如,在噪音污染嚴重的城市環境中,依賴振動通訊的蜘蛛可能需要調整其信號的頻率或強度以確保有效通訊。同樣,光污染可能影響依賴視覺信號的蜘蛛物種(種)的通訊效率。
研究蜘蛛的通訊方式不僅有助於我們理解這些生物的行為和生態,還可能為生物仿生學提供靈感。例如,蜘蛛的振動通訊系統可能為開發新型傳感器或通訊設備提供思路。
7.5 蜘蛛的特殊行為
蜘蛛是自然界中一群極其獨特的生物,它們展現出許多令人驚嘆的特殊行為。這些行為不僅反映了蜘蛛在漫長進化過程中的適應性,也展現了它們在生存和繁衍策略上的創新。
其中最引人注目的特殊行為之一是蜘蛛的模仿行為。許多蜘蛛種類已經進化出驚人的偽裝能力,能夠模仿周圍環境中的其他生物或物體。例如,蘭花蛛(Thomisus onustus)不僅外形酷似花朵,還能根據所處的花朵改變自身的顏色,從而完美地融入環境中。這種高度專業化的擬態不僅能幫助蜘蛛躲避天敵,還能讓它們更容易捕獲獵物。另一個著名的例子是螞蟻蛛(Myrmarachne formicaria),它們不僅在外形上酷似螞蟻,連行走方式都模仿得惟妙惟肖,這種行為不僅能夠保護自己,還能讓它們更容易接近獵物。
一些蜘蛛種類展現出令人驚訝的社會性行為,這在節肢動物中相對罕見。例如,非洲社會性蜘蛛(Stegodyphus dumicola)會共同織網、分享食物,甚至合作撫養幼蛛。這種高度社會化的行為讓它們能夠捕獲比自身大得多的獵物,並在惡劣環境中更好地生存。這種行為挑戰了我們對蜘蛛作為獨居動物的傳統認知。
蜘蛛的氣球飛行行為也是一種極為獨特的適應。許多小型蜘蛛,特別是幼蛛,會分泌出一種特殊的絲,在微風的作用下被帶到空中,從而實現長距離散布。這種行為被稱為"氣球飛行",能夠幫助蜘蛛快速擴散到新的棲息地。有趣的是,科學家發現一些蜘蛛甚至能夠利用地球的電場來增強這種飛行能力,這種行為在動物界中是極為罕見的。
某些蜘蛛種類展現出驚人的建築能力。例如,吊篋蛛(Argyroneta aquatica)是已知唯一能夠在水下生活的蜘蛛。它們會在水下建造一個充滿空氣的"潛水鐘",這個結構不僅作為它們的住所,還作為捕獵和繁殖的場所。這種獨特的適應讓吊篋蛛能夠佔據一個幾乎沒有競爭者的生態位。
在捕獵行為上,某些蜘蛛種類也表現出非常特殊的策略。例如,捕鳥蛛(Theraphosidae)不僅能夠捕食昆蟲,還能捕獲小型脊椎動物如青蛙、蜥蜴,甚至小型鳥類。這種行為在無脊椎動物中極為罕見,展示了蜘蛛驚人的適應能力和生態重要性。
一些蜘蛛種類還展現出複雜的學習能力。研究發現,跳蛛(Salticidae)不僅具有優秀的視力,還能夠學習和記憶複雜的迷宮結構,這種認知能力在節肢動物中是相當罕見的。這表明即使是相對簡單的神經系統,也能夠支持複雜的學習和決策過程。
在繁殖行為方面,某些蜘蛛種類表現出極為獨特的策略。例如,雄性絲蛛(Pisaurina mira)會在交配時將自己的身體獻祭給雌性,這種極端的交配行為被稱為性食同類。雖然這種行為看似不利於個體生存,但實際上能夠大大提高後代的存活率,是一種高度進化的繁殖策略。
蜘蛛的網絡建構行為也展現出令人驚嘆的複雜性和適應性。某些蜘蛛能夠根據環境條件和獵物類型調整其網絡結構。例如,金園蛛(Nephila clavipes)能夠根據風力的強弱來調整蛛網的彈性和黏性,以確保網絡在各種天氣條件下都能有效捕獲獵物。
這些特殊行為不僅展示了蜘蛛驚人的適應能力,也凸顯了它們在生態系統中的重要地位。蜘蛛通過這些獨特的行為,在捕食、繁衍、散布等方面都表現出極強的競爭力,使得它們能夠在各種複雜的生態環境中繁衍生息。同時,這些行為也為我們提供了研究動物行為、生態適應以及進化過程的寶貴素材。隨著研究技術的不斷進步,相信我們會發現更多蜘蛛的特殊行為,進一步揭示這些神奇生物的奧秘。
8. 蜘蛛的繁殖與生活史
蜘蛛的繁殖與生活史是一個充滿奇妙和多樣性的過程。大多數蜘蛛種類都是卵生的,雌性蜘蛛在交配後會產下卵囊。這些卵囊通常由絲質包裹,能夠保護蛛卵免受捕食者和惡劣環境的侵害。卵囊的大小、形狀和數量因蜘蛛種類而異,有些蜘蛛可能一次產下數百個卵。
蜘蛛的生活史通常包括卵、幼蛛和成蛛三個階段。幼蛛孵化後通常會經歷多次蛻皮才能成長為成蛛。有趣的是,某些蜘蛛種類的幼蛛會有群居行為,但隨著成長逐漸變得獨居。例如,園蛛(Araneidae)的幼蛛初期會聚集在一起,共同織網捕食,但隨著體型增大,它們會分散並建立各自的領地。
在繁殖策略方面,蜘蛛展現出驚人的多樣性。有些蜘蛛種類的雄性會在交配時將自己獻祭給雌性,成為雌性的食物,這種行為被稱為性食同類。例如,澳洲紅背蜘蛛(Latrodectus hasselti)的雄性在交配過程中會主動將腹部送入雌性口中。這種看似自殺的行為實際上能夠延長交配時間,提高授精成功率,並為後代提供額外的營養。
某些蜘蛛種類還表現出複雜的親代照顧行為。例如,狼蛛(Lycosidae)的雌性會將卵囊附在紡絲器上隨身攜帶,直到幼蛛孵化。孵化後的幼蛛會爬到母親的背上,由母親照顧和保護一段時間。這種行為大大提高了後代的存活率。
蜘蛛的生活史長短差異很大,有些小型蜘蛛可能在幾個月內完成整個生命週期,而大型蜘蛛如捕鳥蛛(Theraphosidae)可能需要多年才能達到性成熟。這種生活史策略的多樣性反映了蜘蛛對不同環境的適應,也是它們在自然界中廣泛分布的重要原因之一。
8.1 蜘蛛的求偶行為
蜘蛛的求偶行為是一個複雜而引人入勝的過程,展現了自然界中最為精巧和多樣化的行為模式之一。這種行為不僅涉及複雜的交流方式,還包含了精心設計的策略,以確保繁衍成功並避免被誤認為獵物而遭到攻擊。
在大多數蜘蛛種類中,求偶過程通常由雄性主動發起。雄性蜘蛛需要小心謹慎地接近雌性,因為在許多情況下,體型較大的雌性可能會將接近的雄性誤認為獵物。為了避免這種危險,雄性蜘蛛進化出了一系列獨特的求偶行為和信號。
一種常見的求偶策略是振動通訊。許多織網蜘蛛,如園蛛科(Araneidae)的成員,會通過特定的方式拉動雌性的網絲來傳遞求偶信號。這種振動模式與捕獲獵物時產生的振動明顯不同,能夠讓雌性識別出這是一個潛在的配偶而非獵物。例如,金園蛛(Nephila clavipes)的雄性會在雌性網絲上產生一種特定頻率的振動,這種振動能夠有效地吸引雌性的注意並表明自己的存在。
視覺信號在某些蜘蛛種類的求偶行為中扮演著重要角色,尤其是在跳蛛科(Salticidae)中。跳蛛擁有節肢動物中最為發達的視覺系統,能夠感知細微的動作和顏色變化。雄性跳蛛會展示複雜的求偶舞蹈,包括特定的肢體動作、身體擺動,以及鮮艷體色的展示。例如,孔雀跳蛛(Maratus volans)的雄性擁有一個色彩斑斕的腹部翼片,在求偶時會像孔雀開屏一樣展開,並配合特定的舞蹈動作來吸引雌性。這種視覺表演不僅展示了雄性的健康狀況和基因品質,還能幫助雌性識別出同種的雄性。
化學通訊也是蜘蛛求偶行為中的重要組成部分。許多蜘蛛種類會分泌特殊的信息素來吸引異性。雌性蜘蛛通常會在蛛網或周圍環境中留下化學信號,雄性蜘蛛能夠通過感知這些信號來定位潛在的配偶。例如,在狼蛛科(Lycosidae)中,雌性會分泌一種特殊的信息素,雄性能夠通過感知這種化學信號來追蹤雌性的行蹤。
某些蜘蛛種類在求偶過程中還會使用"禮物"策略。雄性會為雌性帶來食物作為求偶禮物,這不僅可以減少被雌性攻擊的風險,還能增加交配的機會。例如,長頜蛛科(Tetragnathidae)的某些種類中,雄性會捕獲一隻昆蟲,用絲包裹後帶給雌性作為求偶禮物。這種行為不僅展示了雄性的捕獵能力,還能為雌性提供額外的營養,有利於後代的發展。
在一些蜘蛛種類中,雄性會採取極端的策略來增加交配成功率。例如,某些球蛛(Araneidae)的雄性會在交配過程中主動斷掉自己的交配器,留在雌性體內。這種行為被稱為"交配栓塞",可以阻止其他雄性與該雌性交配,從而確保自己的精子能夠成功受精。
蜘蛛的求偶行為還涉及複雜的時間和空間選擇。某些種類的雄性會在雌性蛻皮後立即展開求偶,因為這時的雌性往往更容易接受交配。例如,寡婦蛛屬(Latrodectus)的雄性會在雌性即將完成最後一次蛻皮時就開始守候在旁,以便在最佳時機展開求偶。
在一些社會性蜘蛛種類中,求偶行為可能涉及更為複雜的群體互動。例如,在非洲社會性蜘蛛(Stegodyphus dumicola)中,多個雄性可能同時在一個雌性周圍展開求偶,形成一種競爭性的求偶場景。這種情況下,雌性的選擇不僅基於個體的求偶表現,還可能受到群體動態的影響。
值得注意的是,蜘蛛的求偶行為還需要克服巨大的體型差異帶來的挑戰。在許多蜘蛛種類中,雌性的體型遠大於雄性,這使得求偶過程充滿危險。為了應對這一挑戰,一些蜘蛛種類演化出了特殊的策略。例如,某些蠅虎蛛(Thomisidae)的雄性會在雌性捕食時悄悄接近並快速完成交配。
蜘蛛的求偶行為不僅展示了自然選擇的力量,也反映了性選擇在塑造這些行為中的重要作用。這些複雜多樣的求偶策略確保了基因的有效傳遞,同時也為我們提供了研究動物行為演化的寶貴素材。隨著研究技術的不斷進步,科學家們正在揭示更多關於蜘蛛求偶行為的奧秘,加深我們對這一迷人領域的理解。
8.2 蜘蛛的交配系統
蜘蛛的交配系統展現了驚人的多樣性和複雜性,反映了它們在不同生態環境下的適應策略。這些交配系統不僅涉及生理和行為上的適應,還包括了複雜的生態和進化因素。
一種常見的蜘蛛交配系統是多配偶制。在這種系統中,雌性和雄性都可能與多個異性交配。這種策略可以增加遺傳多樣性,提高後代的適應能力。例如,在園蛛科(Araneidae)中,雌性通常會與多個雄性交配。這不僅可以確保充足的精子供應,還能夠通過精子競爭來選擇品質最好的精子。雄性園蛛則會試圖與盡可能多的雌性交配,以最大化其繁殖成功率。
然而,某些蜘蛛種類則採用單配偶制。在這種系統中,雌性和雄性在一個繁殖季節只與一個配偶交配。這種策略通常出現在資源稀缺或繁殖機會有限的環境中。例如,某些隱蛛科(Clubionidae)的成員就採用這種策略。雄性隱蛛會在雌性成熟之前就開始守候,並在雌性完成最後一次蛻皮後立即交配。這種行為可以確保雄性成為雌性的唯一交配對象,從而最大化其繁殖成功率。
一些蜘蛛種類則展現出更為極端的交配系統,如雌性專一制。在這種系統中,雌性終生只與一個雄性交配,而雄性則可能與多個雌性交配。這種策略通常與雄性的自我犧牲行為相關聯。例如,在澳洲紅背蜘蛛(Latrodectus hasselti)中,雄性在交配過程中會將自己的腹部送入雌性口中,成為雌性的食物。這種行為不僅延長了交配時間,增加了受精的機會,還為雌性提供了額外的營養,有利於後代的發展。
相反地,某些蜘蛛種類則表現出雄性專一制。在這種系統中,雄性終生只與一個雌性交配,而雌性可能與多個雄性交配。這種策略通常出現在雄性投資較大的情況下。例如,在某些結網蠅虎蛛(Dictynidae)中,雄性會在雌性的網上建立自己的小網,並長期居住在那裡,幫助雌性維護網絡和照顧後代。
蜘蛛的交配系統還包括一些獨特的變體,如序列多偶制。在這種系統中,蜘蛛在不同的時期可能採用不同的交配策略。例如,某些狼蛛科(Lycosidae)的成員在繁殖季節初期可能採用多配偶制,但隨著季節推進,可能轉向單配偶制。這種靈活的策略允許蜘蛛根據環境條件和個體狀況調整其繁殖策略。
另一種有趣的交配系統是所謂的"交配競賽"。在這種系統中,多個雄性同時在一個雌性周圍展開激烈的競爭。這種情況常見於社會性蜘蛛或者雌性聚集的地方。例如,在某些金蛛科(Nephilidae)的成員中,多個體型較小的雄性會聚集在一個大型雌性的網上,彼此競爭交配機會。這種競爭不僅涉及直接的身體衝突,還包括複雜的行為策略,如守衛、干擾其他雄性的求偶等。
蜘蛛的交配系統還常常與特殊的形態學特徵相關聯。例如,在某些蛛科(Theriidae)中,雄性的交配器官在交配後會斷裂,留在雌性體內形成"交配栓"。這種策略可以防止其他雄性的精子進入,確保自己的基因得以傳播。然而,這也意味著雄性只能交配一次,這就導致了極為激烈的交配競爭。
值得注意的是,蜘蛛的交配系統還受到環境因素的強烈影響。例如,在資源豐富的環境中,多配偶制可能更為普遍,因為個體有足夠的能量支持多次交配和產卵。相反,在資源稀缺的環境中,單配偶制可能更有優勢,因為它可以最大化有限資源的利用效率。
此外,蜘蛛的交配系統還與其生活史特徵密切相關。例如,壽命較短的蜘蛛種類可能更傾向於採用多配偶制,以在有限的時間內最大化繁殖機會。而壽命較長的種類則可能採用更為保守的策略,如單配偶制或序列多偶制。
蜘蛛的交配系統研究不僅對於理解蜘蛛的生態和進化具有重要意義,還為我們提供了研究性選擇、資源分配和行為生態學的寶貴模型。通過深入研究這些複雜的交配系統,科學家們不僅能夠更好地理解蜘蛛的生活史策略,還能夠為更廣泛的生態學和進化生物學問題提供新的洞見。
8.3 性選擇與性二型性
蜘蛛的世界中,性選擇和性二型性是兩個緊密相連且極為引人入勝的現象。這些現象不僅反映了蜘蛛的進化歷程,還展示了自然選擇如何塑造生物的形態和行為。
性選擇是達爾文提出的一種特殊的自然選擇形式,它主要關注那些影響繁殖成功的特徵。在蜘蛛中,性選擇通常表現為雌性對雄性特徵的選擇,或者雄性之間為爭奪交配機會而產生的競爭。這種選擇壓力往往導致雌雄之間在形態、大小、顏色甚至行為上的顯著差異,也就是我們所說的性二型性。
蜘蛛的性二型性在許多方面都表現得非常明顯。最常見的是體型差異,在大多數蜘蛛種類中,雌性的體型都明顯大於雄性。這種差異在某些種類中尤為突出,例如在金園蛛(Nephila clavipes)中,雌性的體長可以達到雄性的五倍以上。這種巨大的體型差異可能源於多重因素的共同作用。首先,較大的體型使雌性能夠產生更多的卵,從而提高繁殖成功率。其次,在許多蜘蛛種類中,雌性需要攜帶和保護卵囊,較大的體型有利於這一任務的完成。此外,較大的體型也使雌性在捕獵時具有優勢,能夠獲取更多的食物資源。
然而,雄性蜘蛛的小體型也並非沒有優勢。在許多情況下,較小的體型使雄性能夠更快速、更靈活地移動,這在尋找雌性和逃避被捕食時都是有利的。例如,在跳蛛科(Salticidae)中,雄性通常比雌性小,但卻擁有更長的腿,這使它們能夠更快速地移動,有利於在複雜的環境中搜尋配偶。
除了體型差異,顏色和形態上的性二型性在蜘蛛中也很常見。在許多種類中,雄性往往比雌性更為艷麗或擁有更為複雜的體表紋路。這種現象在跳蛛中尤為明顯,如孔雀跳蛛(Maratus volans)的雄性擁有色彩斑斕的腹部翼片,用於求偶展示。這種鮮艷的色彩是性選擇的結果,雌性傾向於選擇那些顏色更鮮艷、展示更出色的雄性作為配偶。然而,這種艷麗的外表也可能增加被捕食的風險,這就形成了性選擇和自然選擇之間的權衡。
在某些蜘蛛種類中,雄性還進化出了特殊的形態結構用於求偶或者與其他雄性競爭。例如,某些皿蛛科(Pisauridae)的雄性擁有特化的前足,用於在求偶時抓住雌性。這種特化的結構增加了雄性在競爭中的優勢,但也可能影響其在其他方面的表現,如捕獵能力。
蜘蛛的性選擇和性二型性還表現在行為方面。在許多種類中,雄性表現出更為積極的求偶行為,而雌性則更為被動。這種行為差異可能是由於雌性在繁殖中投入更多,因此更為謹慎地選擇配偶。例如,在園蛛科(Araneidae)中,雄性會主動尋找雌性的網並進行求偶,而雌性則會仔細評估雄性的求偶表現再決定是否接受交配。
性選擇還可能導致蜘蛛演化出一些極端的策略。例如,在某些球蛛(Araneidae)中,雄性會在交配過程中主動斷掉自己的交配器,留在雌性體內形成"交配栓"。這種行為雖然使雄性失去了再次交配的機會,但卻能有效防止其他雄性的精子進入雌性體內,從而提高自己的繁殖成功率。這種極端的策略反映了性選擇的強大力量。
另一個有趣的現象是某些蜘蛛種類中存在的雄性二型性。例如,在某些金蛛科(Nephilidae)的成員中,同一種蜘蛛的雄性可能存在兩種不同的形態:一種是體型較大、更具攻擊性的"主導型"雄性,另一種是體型較小、更為靈活的"潛行型"雄性。這兩種類型採用不同的策略來獲得交配機會,反映了複雜的性選擇壓力。
蜘蛛的性選擇和性二型性還與其生態環境密切相關。例如,在資源稀缺的環境中,雌性可能更傾向於選擇那些能夠提供"求偶禮物"的雄性。這種禮物通常是被絲包裹的獵物,不僅可以減少雌性對雄性的攻擊性,還能為雌性提供額外的營養。這種行為在長頜蛛科(Tetragnathidae)的某些成員中就有觀察到。
研究蜘蛛的性選擇和性二型性不僅有助於我們理解蜘蛛的進化歷程,還為我們提供了研究性選擇理論的絕佳模型。這些研究可以幫助我們更好地理解生物如何在繁殖成功和生存之間取得平衡,以及性選擇如何驅動物種的多樣化。隨著研究方法的不斷進步,我們對蜘蛛性選擇和性二型性的理解也在不斷深化,為進化生物學和行為生態學提供了豐富的研究素材。
8.4 蜘蛛的性食同類現象
蜘蛛的性食同類現象是自然界中最為引人注目且具爭議性的行為之一。這種行為指的是在交配過程中或交配之後,雌性蜘蛛捕食並吞噬雄性蜘蛛的現象。儘管這種行為在人類看來可能顯得殘酷,但在蜘蛛的世界中,它是一種複雜的進化策略,涉及多方面的生態和進化因素。
性食同類現象在多個蜘蛛科中都有發現,但以黑寡婦蛛屬(Latrodectus)和狼蛛科(Lycosidae)中最為常見和極端。以澳洲紅背蜘蛛(Latrodectus hasselti)為例,雄性在交配過程中會主動將自己的腹部送入雌性口中。這種看似自殺的行為實際上是一種精心設計的繁殖策略。當雌性開始進食時,雄性可以延長交配時間,增加精子傳遞的機會。更重要的是,被吃掉的雄性為雌性提供了豐富的營養,這些營養可以直接轉化為更多更健康的後代。
然而,並非所有的性食同類行為都是雄性主動犧牲的結果。在許多情況下,雌性會在交配結束後突然攻擊並吞噬雄性。這種行為可能源於多個原因。首先,蜘蛛是典型的獨居捕食者,對任何接近的生物都有攻擊傾向。其次,雌性通常比雄性大得多,這使得雄性成為易於捕獲的獵物。此外,交配後的雌性需要大量營養來支持卵子的發育,而雄性恰好是一個便利的營養來源。
從進化的角度來看,性食同類現象可能為雌性蜘蛛帶來多重好處。首先,它提供了額外的營養來源,有助於產生更多或更健康的後代。研究表明,被吃掉的雄性可以為雌性提供高達30%的額外營養。其次,這種行為可以確保雌性只與一個雄性交配,從而避免了多次交配可能帶來的風險,如受傷或感染。最後,它可能是一種選擇機制,只有最強壯、最敏捷的雄性才能在交配後逃脫,從而確保了優質基因的傳遞。
對於雄性蜘蛛來說,性食同類現象無疑是一個巨大的風險。然而,從進化的角度來看,如果被吃掉能夠顯著增加後代的數量或質量,那麼這種自我犧牲的行為也是有利的。一些研究發現,在某些蜘蛛種類中,被吃掉的雄性比成功逃脫的雄性產生更多的後代。這是因為雌性可能更傾向於使用被吃掉的雄性的精子來受精。
值得注意的是,性食同類現象並非在所有蜘蛛種類中都普遍存在,其發生頻率和嚴重程度也因種類而異。例如,在某些園蛛科(Araneidae)的成員中,性食同類現象相對罕見,雄性通常能夠在交配後安全離開。相比之下,在某些姬蛛科(Theridiidae)的成員中,性食同類現象則非常普遍。
蜘蛛的性食同類行為還受到多種環境因素的影響。例如,食物的可用性就是一個重要因素。研究發現,當環境中食物稀缺時,性食同類現象的發生率會增加。這可能是因為在資源匱乏的情況下,雌性更需要額外的營養來支持卵子的發育。另一個影響因素是雌性的交配歷史。一些研究表明,已經交配過的雌性比處女雌性更容易表現出性食同類行為,這可能是因為已交配的雌性不再那麼迫切需要精子。
雄性蜘蛛也進化出了多種策略來應對性食同類的風險。一些雄性會在交配前為雌性帶來"求偶禮物",通常是被絲包裹的獵物。這不僅可以分散雌性的注意力,還能為其提供額外的營養,從而降低被吃掉的風險。另一種策略是選擇最佳的交配時機。例如,一些雄性會在雌性剛剛進食後或正在蛻皮時接近,因為這時雌性不太可能發動攻擊。
有趣的是,一些雄性蜘蛛甚至進化出了特殊的形態或行為來應對性食同類的威脅。例如,某些蠅虎蛛科(Thomisidae)的雄性擁有特別長的前腿,可以在交配時將雌性抱住,防止其發動攻擊。還有一些雄性會在交配後迅速逃離,或者採取特殊的姿勢來保護自己的重要部位。
蜘蛛的性食同類現象為我們提供了研究性選擇、親代投資和行為生態學的寶貴模型。這種行為挑戰了我們對繁殖策略的傳統理解,展示了自然選擇如何在看似矛盾的行為中找到平衡。通過深入研究這一現象,科學家們不僅能夠更好地理解蜘蛛的生態和進化,還能為更廣泛的生物學問題提供新的洞見。
8.5 蜘蛛的親代照顧
蜘蛛的親代照顧行為是一個引人入勝且多樣化的話題,展現了這些八腳動物驚人的適應能力和進化策略。雖然人們常常認為蜘蛛是冷血無情的生物,但實際上,許多蜘蛛種類表現出令人驚訝的親代投資和照顧行為。
在大多數蜘蛛種類中,親代照顧主要由雌性承擔。這種照顧通常始於產卵階段,雌性蜘蛛會精心選擇產卵地點,並為卵囊提供保護。例如,狼蛛科(Lycosidae)的雌性會將卵囊附著在自己的紡絲器上,隨身攜帶。這種行為不僅可以保護卵囊免受捕食者和寄生蟲的侵害,還能確保卵囊處於最佳的溫度和濕度環境中。狼蛛媽媽會定期將卵囊翻轉,確保所有的卵都能均勻發育。如果卵囊不慎掉落,母蛛會立即返回尋找並重新附著。
某些蜘蛛種類的親代照顧甚至延續到幼蛛孵化之後。以狼蛛為例,幼蛛孵化後會爬到母蛛的背上,由母蛛攜帶和保護一段時間。這期間,幼蛛依靠卵黃囊生存,不需要額外餵食。當幼蛛完成第一次蛻皮後,它們會逐漸離開母親,開始獨立生活。這種延長的親代照顧大大提高了幼蛛的存活率,特別是在資源匱乏或環境惡劣的情況下。
在一些織網蜘蛛種類中,我們可以觀察到更為複雜的親代照顧行為。例如,某些園蛛科(Araneidae)的成員會在自己的大網旁邊為卵囊織一個特殊的保護網。母蛛會在這個保護網附近守衛,直到幼蛛孵化。有趣的是,一些種類的母蛛甚至會為初生的幼蛛預先織好小型捕獵網,幫助它們開始獨立生活。
某些蜘蛛種類表現出更為極端的親代照顧行為。例如,智利巨型捕鳥蛛(Grammostola rosea)的雌性會在孵化期間絕食長達兩個月,專注於保護和照顧卵囊。這種行為雖然對母蛛的生存構成挑戰,但可以大大提高後代的存活率。
在一些社會性蜘蛛種類中,親代照顧行為更是達到了令人驚嘆的程度。例如,非洲社會性蛛(Stegodyphus dumicola)不僅共同織網、捕獵,還會合作撫養幼蛛。在這個物種中,無子女的雌性會幫助照顧其他個體的後代,這種行為在節肢動物中相當罕見,更接近於某些哺乳動物和鳥類的社會結構。
有趣的是,某些蜘蛛種類的雌性會通過自我犧牲來為後代提供營養。這種行為被稱為"母體餵養"(Matriphagy)。例如,在某些姬蛛科(Theridiidae)的成員中,幼蛛會在孵化後不久開始吞噬自己的母親。這種極端的親代投資確保了幼蛛在生命初期獲得充足的營養,大大提高了它們的存活率。雖然這種行為從人類的角度看似殘酷,但從進化的角度來看,如果這種自我犧牲能夠顯著提高後代的存活率和適應度,那麼它就是一種有效的繁殖策略。
值得注意的是,雖然親代照顧在蜘蛛中主要由雌性承擔,但在某些種類中,雄性也會參與其中。例如,在某些盜蛛科(Pisauridae)的成員中,雄性會幫助守護卵囊和照顧幼蛛。這種行為可能是為了增加自己基因傳遞的機會,特別是在一夫一妻制的物種中。
蜘蛛的親代照顧行為還會受到環境因素的影響。例如,在食物資源豐富的環境中,親代投資可能會增加,因為母蛛有足夠的能量來支持延長的照顧期。相反,在資源匱乏的環境中,母蛛可能會減少投資,將更多精力用於自身的生存。
氣候條件也會影響親代照顧的程度和方式。在寒冷或乾燥的環境中,某些蜘蛛種類會為卵囊提供額外的保護。例如,某些跳蛛科(Salticidae)的成員會在寒冷季節將卵囊藏在樹皮下或岩石縫隙中,並用特殊的絲覆蓋,以提供額外的保溫和防潮效果。
蜘蛛的親代照顧行為不僅對個體的繁殖成功至關重要,還對整個種群的動態產生深遠影響。那些提供較多親代照顧的種類通常產卵量較少,但幼體的存活率較高。這種策略在穩定或資源有限的環境中可能更有優勢。相反,那些親代照顧較少的種類往往產卵量大,但幼體死亡率也高,這種策略在不穩定或資源豐富的環境中可能更有效。
研究蜘蛛的親代照顧行為不僅有助於我們理解這些神奇生物的生活史策略,還為研究動物行為學、進化生物學和生態學提供了寶貴的見解。通過比較不同蜘蛛種類的親代照顧策略,科學家們可以更好地理解親代投資如何隨環境條件和進化壓力而變化。這些研究還可以為保護瀕危蜘蛛種類提供重要信息,幫助制定更有效的保護策略。
8.6 蜘蛛的生活週期
蜘蛛的生活週期是一個引人入勝的過程,展現了這些八腳動物驚人的適應能力和生存策略。從卵到成熟個體,蜘蛛經歷了一系列複雜的發育階段,每個階段都有其獨特的特徵和挑戰。
蜘蛛的生命旅程始於卵階段。雌性蜘蛛通常會將卵產在精心編織的絲質卵囊中。這些卵囊不僅能保護卵免受捕食者和環境因素的侵害,還能維持適宜的溫度和濕度。卵囊的大小、形狀和數量因蜘蛛種類而異。例如,園蛛科(Araneidae)的某些成員可能一次產下數百個卵,而捕鳥蛛科(Theraphosidae)的成員則可能只產下幾十個卵。
在適宜的條件下,卵會在卵囊內孵化。孵化的時間因種類和環境條件而異,可能短至幾天,也可能長達數月。例如,在溫帶地區,某些蜘蛛種類的卵會在秋季產下,但要等到來年春天才會孵化。這種延遲孵化的策略可以幫助幼蛛避開不利的冬季條件。
孵化後的幼蛛通常會經歷幾次蛻皮才能達到成熟階段。這個過程被稱為"不完全變態",因為幼蛛的外形已經與成體相似,只是體型較小。第一次蛻皮通常在卵囊內完成,這個階段的幼蛛被稱為前若蟲(Postembryo)。這個階段的幼蛛通常不會進食,而是依靠卵黃提供的營養生存。
離開卵囊後,幼蛛進入若蟲階段。這個時期,幼蛛開始獨立捕食和生活。許多種類的幼蛛會通過"氣球飄行"(Ballooning)來擴散。它們會爬到高處,釋放出一束絲線,藉助風力飄到新的棲息地。這種行為使得蜘蛛能夠迅速擴散到新的區域,是它們適應能力強的重要原因之一。
在成長過程中,蜘蛛會經歷多次蛻皮。蛻皮是蜘蛛生命中的關鍵時刻,也是最脆弱的時期。在蛻皮過程中,蜘蛛會停止進食,靜止不動,等待新的外骨骼硬化。這個過程可能持續數小時到數天不等。蛻皮的次數因種類而異,小型蜘蛛可能只需要4-5次蛻皮就能達到成熟,而大型蜘蛛如捕鳥蛛可能需要10次以上的蛻皮。
值得注意的是,蜘蛛的生長過程中存在性別差異。在許多種類中,雄性比雌性更快達到成熟階段,並且蛻皮次數較少。這種策略可能是為了盡快進入繁殖階段,增加找到雌性和交配的機會。
當蜘蛛達到性成熟後,它們就進入了成體階段。這個階段的主要任務是繁衍後代。雄性蜘蛛會積極尋找雌性,進行求偶和交配。有趣的是,某些蜘蛛種類的雄性會在最後一次蛻皮時發育出特殊的交配器官,稱為雄蛛須肢(Pedipalps)。這些器官用於將精子傳遞給雌性。
成體蜘蛛的壽命因種類而異。小型蜘蛛的壽命可能只有幾個月,而大型蜘蛛如某些捕鳥蛛種類可能活20年以上。在野外,蜘蛛的實際壽命往往受到多種因素的影響,如捕食、疾病、環境條件等。
蜘蛛的生活週期還會受到季節變化的影響。在溫帶地區,許多蜘蛛種類會採取特殊的策略來度過寒冷的冬季。有些種類會以成體或亞成體的形態過冬,躲在樹皮下或其他庇護所中。另一些種類則會以卵的形態過冬,來年春天才孵化。例如,交織貓蛛(Araneus diadematus)通常在秋季產卵,卵在冬季保持休眠狀態,直到春季才孵化。
某些蜘蛛種類展現出更為特殊的生活週期。例如,澳洲紅背蛛(Latrodectus hasselti)的雄性在交配後往往會被雌性吃掉。這種被稱為"性食同類"的行為雖然看似殘酷,但從進化的角度來看,可能有利於後代的生存,因為被吃掉的雄性為雌性提供了額外的營養來源。
蜘蛛的生活週期還與其生態位密切相關。例如,織網蜘蛛和遊獵蜘蛛可能有不同的生活史策略。織網蜘蛛通常有較穩定的棲息地,可能有更長的生命週期和更多的繁殖機會。相比之下,遊獵蜘蛛可能需要更頻繁地移動,因此可能有較短的生命週期但更高的繁殖率。
理解蜘蛛的生活週期對於生態學研究和保護工作都具有重要意義。通過研究不同階段的生存率、發育時間和繁殖策略,科學家們可以更好地理解蜘蛛種群的動態變化和生態功能。這些知識不僅有助於我們保護瀕危的蜘蛛種類,還能為生態系統管理提供重要參考。例如,了解農田中蜘蛛的生活週期可以幫助農民更好地利用這些天然的害蟲控制者,減少農藥的使用。
9. 蜘蛛的覓食生態
蜘蛛的覓食生態是一個豐富多樣的領域,反映了這些捕食者在自然界中的重要地位。作為廣泛分布的節肢動物捕食者,蜘蛛在控制昆蟲種群和維持生態平衡方面發揮著關鍵作用。
蜘蛛主要以昆蟲和其他小型節肢動物為食,但某些大型種類如捕鳥蛛甚至能捕食小型脊椎動物。蜘蛛的覓食策略可大致分為兩類:織網捕獵和主動獵捕。織網蜘蛛如園蛛科(Araneidae)的成員會編織精巧的網來捕獲飛行昆蟲,而遊獵蜘蛛如狼蛛科(Lycosidae)的成員則主動追捕獵物。
蜘蛛的捕獵過程通常涉及毒液的使用。當獵物被捕獲後,蜘蛛會注射毒液以麻痺或殺死獵物。隨後,蜘蛛會分泌消化酶,將獵物體內組織液化,然後吸食這些液體。這種外部消化的方式使蜘蛛能夠捕食比自身大得多的獵物。
蜘蛛的食性廣泛,但也存在一定的選擇性。例如,某些蜘蛛種類專門捕食螞蟻,而另一些則偏好飛蛾。這種食性的專一化反映了蜘蛛與其獵物之間的協同進化關係。
值得注意的是,蜘蛛的覓食行為還受到環境因素的影響。例如,在食物稀缺的時期,蜘蛛可能會變得更具侵略性,或者調整其網的結構以提高捕獲效率。這種靈活的覓食策略使蜘蛛能夠在各種環境中生存和繁衍,從而在生態系統中扮演著重要角色。
9.1 蜘蛛的捕獵技巧
蜘蛛的捕獵技巧是自然界中最為精妙和多樣化的生存策略之一。這些八腳獵手經過數億年的進化,發展出了一系列令人驚嘆的捕獵方法,使它們能夠在各種環境中成功捕獲獵物。
最為人熟知的蜘蛛捕獵技巧莫過於織網。織網蜘蛛,如園蛛科(Araneidae)的成員,能夠編織出複雜而精巧的捕獵網。這些網不僅是捕獲獵物的工具,更是蜘蛛感知周圍環境的延伸。當昆蟲撞上蛛網時,振動會迅速傳遞到蜘蛛的腳部,蜘蛛能夠通過這些振動判斷獵物的大小、位置甚至類型。例如,金園蛛(Nephila clavipes)能夠根據不同獵物撞擊網絲時產生的特定振動頻率來區分蝴蝶和蜜蜂。
有趣的是,不同種類的蜘蛛會織造不同類型的網。除了常見的圓形網外,還有漏斗狀網、片狀網等。每種網的結構都適應於捕捉特定類型的獵物。例如,漏斗網蛛(Agelenidae)織造的漏斗狀網特別適合捕捉在地面活動的昆蟲,而片狀網則更適合捕捉飛行昆蟲。
然而,並非所有蜘蛛都依賴蛛網捕獵。許多蜘蛛採用主動獵捕的策略。狼蛛科(Lycosidae)的成員就是典型的遊獵蜘蛛,它們靠敏銳的視力和快速的反應能力來追捕獵物。這些蜘蛛通常在地面上活動,能夠迅速發現並追趕獵物。有些狼蛛甚至能夠在水面上奔跑,捕捉水生昆蟲。
跳蛛科(Salticidae)的成員則展示了另一種精湛的主動捕獵技巧。這些蜘蛛擁有節肢動物中最發達的視覺系統,能夠精確判斷獵物的距離和方向。它們會小心翼翼地接近獵物,然後突然跳躍攻擊。跳蛛的跳躍能力非常驚人,有些種類能夠跳躍超過自身體長50倍的距離。
某些蜘蛛種類還發展出了特殊的擬態捕獵技巧。例如,蟹蛛科(Thomisidae)的成員常常隱藏在花朵上,它們的體色能夠與花瓣完美融合。當昆蟲前來採蜜時,這些蜘蛛就會突然出擊。澳大利亞的一種稀有蜘蛛,鳥糞蛛(Celaenia excavata)則更為特別。它的外形和氣味都酷似鳥類排泄物,能夠吸引以糞便為食的昆蟲靠近,然後將其捕獲。
有些蜘蛛還開發出了使用工具的捕獵技巧。比如,捕網蛛科(Deinopidae)的成員被稱為「網魔蛛」,它們會用前足拉伸一小片特製的黏性網,當獵物接近時,蜘蛛會迅速將這片網拋出,將獵物罩住。這種行為被認為是節肢動物中最接近使用工具的例子之一。
水生蜘蛛的捕獵技巧也十分有趣。例如,泰來洛爾加蛛(Argyroneta aquatica)是已知唯一能夠在水下生活的蜘蛛。它們會在水下植物間建造一個充滿空氣的「潛水鐘」作為居所,並在這個水下基地周圍捕獵小型水生動物。
某些蜘蛛還採用合作捕獵的策略。雖然大多數蜘蛛是獨居的,但有少數社會性蜘蛛種類,如非洲社會性蛛(Stegodyphus dumicola),會共同織網並分享獵物。這種行為使它們能夠捕獲比單獨行動時大得多的獵物。
值得一提的是,蜘蛛的捕獵技巧並非一成不變。許多蜘蛛能夠根據環境條件和獵物類型調整其捕獵策略。例如,當風力較大時,某些織網蜘蛛會增加網絲的彈性,以適應強風條件下的捕獵。
蜘蛛的毒液在捕獵過程中扮演著關鍵角色。大多數蜘蛛的毒液主要用於麻痺或殺死獵物,而非作為防禦工具。有趣的是,某些蜘蛛能夠根據獵物的大小和類型調整注射的毒液量,這種精確的控制能力確保了捕獵的效率和能量的節約。
蜘蛛的這些多樣化和精妙的捕獵技巧不僅體現了自然選擇的力量,也展示了這些生物在生態系統中的重要地位。作為廣泛分布的節肢動物捕食者,蜘蛛在控制昆蟲種群和維持生態平衡方面發揮著不可或缺的作用。通過研究蜘蛛的捕獵技巧,科學家們不僅能夠更好地理解這些神奇生物的行為生態學,還能為生物仿生學和害蟲管理等領域提供寶貴的啟發。
9.2 蜘蛛的食性組成
蜘蛛的食性組成是一個複雜而多樣的主題,反映了這些捕食者在生態系統中的重要地位和適應能力。作為廣泛分布的節肢動物捕食者,蜘蛛的食譜涵蓋了各種小型生物,主要以其他節肢動物為主,但也包括一些更大型的獵物。
蜘蛛的食性主要由其體型、棲息環境和捕獵方式決定。大多數蜘蛛以昆蟲為主要食物來源,這包括蒼蠅、蚊子、蛾子、蝴蝶、甲蟲等。例如,常見的家蛛(Tegenaria domestica)主要捕食家中的小型昆蟲,如蠅類和蚊子。園蛛科(Araneidae)的成員則主要捕捉飛行昆蟲,其圓形網特別適合攔截飛行中的獵物。
然而,蜘蛛的食譜並不僅限於昆蟲。許多蜘蛛也捕食其他節肢動物,如蠍子、蜈蚣、螨蟎等。例如,一些遊獵蜘蛛如狼蛛科(Lycosidae)的成員,會捕食地表活動的各種小型節肢動物。有趣的是,某些專業化的蜘蛛種類甚至以其他蜘蛛為食。這種被稱為「蛛食蛛」的現象在自然界中並不罕見,例如長腳蛛科(Pholcidae)的一些成員就以捕食其他蜘蛛為主。
一些大型蜘蛛種類的食性更為廣泛,甚至包括小型脊椎動物。最著名的例子是捕鳥蛛科(Theraphosidae)的成員,儘管名為「捕鳥」,但它們實際上很少捕食鳥類。然而,它們確實能夠捕獲如小型蜥蜴、青蛙、老鼠等小型脊椎動物。例如,哥斯大黎加斑捕鳥蛛(Megaphobema mesomelas)就曾被觀察到捕食蛇類。
水生或半水生蜘蛛的食性也十分有趣。例如,捕魚蛛(Dolomedes)能夠在水面上行走,它們不僅捕食水生昆蟲,還能捕獲小魚和蝌蚪。泰來洛爾加蛛(Argyroneta aquatica)則完全適應了水下生活,它們在水草間建造充滿空氣的「潛水鐘」,並在周圍捕獵各種小型水生動物。
蜘蛛的食性還會隨季節和生活史階段而變化。在資源豐富的季節,蜘蛛可能會更具選擇性,偏好某些特定類型的獵物。而在食物稀缺的時期,它們可能會變得不那麼挑剔。幼蛛和成蛛的食性也可能有所不同,幼蛛通常捕食較小的獵物,隨著體型增大,它們能夠捕獲更大、更多樣化的獵物。
某些蜘蛛種類展現出高度專業化的食性。例如,蟻蛛科(Zodariidae)的成員專門捕食螞蟻。這些蜘蛛不僅在形態上模仿螞蟻,還能夠抵抗螞蟻的攻擊,這使它們能夠成功地在螞蟻群中獵食。另一個例子是南美洲的吸血蛛(Evarcha culicivora),這種跳蛛偏好捕食已經吸血的蚊子,因為這些蚊子體內含有豐富的營養物質。
蜘蛛的食性組成還受到環境因素的強烈影響。在農業生態系統中,蜘蛛往往成為重要的生物防治工具。它們能夠捕食多種農業害蟲,如蚜蟲、粉蝨等。例如,在水稻田中,狼蛛和蠅虎蛛是控制稻飛蝨等害蟲的重要天敵。
有趣的是,一些蜘蛛種類還表現出食植現象,儘管這在蜘蛛中並不常見。例如,中美洲的巴古錫蛛(Bagheera kiplingi)主要以金合歡樹的葉片為食,這是目前已知唯一一種主要植食性的蜘蛛。這種特殊的食性被認為是對特定生態位的適應。
蜘蛛的食性組成還與其在食物網中的位置密切相關。作為中層捕食者,蜘蛛不僅控制昆蟲等初級消費者的數量,也成為鳥類、蜥蜴等高級捕食者的食物來源。這種複雜的食物網關係使得蜘蛛在生態系統中扮演著關鍵的角色,影響著能量流動和物質循環。
研究蜘蛛的食性組成對於理解生態系統動態和生物多樣性保護具有重要意義。通過分析蜘蛛的食物組成,科學家們可以評估特定環境中的昆蟲多樣性,監測生態系統健康狀況,甚至預測環境變化對生態系統的影響。例如,通過研究城市環境中蜘蛛的食性變化,可以評估城市化對昆蟲群落的影響。
此外,了解蜘蛛的食性組成對於農業害蟲管理也有重要應用。通過鼓勵特定蜘蛛種類在農田中定居,可以有效控制某些農業害蟲的數量,減少農藥的使用。這種生物防治方法不僅環保,還能夠降低農業成本,提高農產品質量。
9.3 毒液在捕獵中的應用
蜘蛛的毒液是一種獨特而複雜的生物武器,在捕獵過程中扮演著至關重要的角色。這種毒液不僅能夠迅速制服獵物,還能輔助消化,使蜘蛛能夠有效地吸收獵物的營養。蜘蛛毒液的成分極為複雜,包含了多種蛋白質、胜肽和其他生物活性物質,這些成分共同作用,產生了多樣化的生理效果。
在捕獵過程中,蜘蛛會通過其特化的螯肢(chelicerae)將毒液注入獵物體內。毒液的主要作用是快速麻痺或殺死獵物,防止其逃脫或反擊。不同種類的蜘蛛,其毒液的組成和作用機制也有所不同,這與它們的獵物類型和捕獵策略密切相關。
例如,有些蜘蛛的毒液主要作用於獵物的神經系統,能夠迅速導致獵物癱瘓。這種策略特別適合捕捉活動能力強、反應迅速的獵物,如飛蟲。另一些蜘蛛的毒液則主要作用於獵物的肌肉組織,造成肌肉麻痺或崩解,這種毒液特別適合捕捉體型較大或有堅硬外骨骼的獵物。
有趣的是,一些蜘蛛物種能夠根據獵物的不同調整其毒液的成分和劑量。這種適應性使得蜘蛛能夠更有效地利用其有限的毒液資源,同時也減少了不必要的能量消耗。例如,澳洲漏斗網蛛(Atrax robustus)就能夠根據獵物的大小和種類來調整其毒液的注射量。
除了直接殺傷獵物外,蜘蛛毒液還具有消化作用。許多蜘蛛物種的毒液含有消化酶,能夠在獵物體內開始進行預消化過程。這種策略特別適合那些體型小、無法吞食大塊獵物的蜘蛛物種。通過注入含有消化酶的毒液,蜘蛛可以將獵物的內部組織液化,然後吸食這些已經預消化的營養物質。
值得注意的是,並非所有蜘蛛都在捕獵過程中大量使用毒液。一些主要依靠蛛網捕獵的蜘蛛物種,如園蛛(Araneidae),通常會先用蛛絲將獵物纏繞固定,然後再視情況使用少量毒液。這種策略既節省了寶貴的毒液資源,又能有效地控制獵物。
蜘蛛毒液在捕獵過程中的應用還體現在其多樣性上。不同種類的蜘蛛可能針對不同類型的獵物演化出特定的毒液成分。例如,專門捕食蟑螂的蜘蛛物種可能擁有特別有效對付蟑螂神經系統的毒素,而專門捕食蝴蝶或飛蛾的蜘蛛則可能擁有更適合麻痺這類昆蟲的毒素。
此外,一些蜘蛛物種還利用毒液來防禦捕食者。雖然大多數蜘蛛的毒液主要用於捕獵,但在面臨威脅時,它們也可能使用毒液作為自衛手段。這種雙重用途的特性使得毒液成為蜘蛛生存競爭中的重要工具。
研究蜘蛛毒液在捕獵中的應用不僅對了解蜘蛛的生態學行為具有重要意義,還為人類在醫學和農業等領域提供了寶貴的靈感。許多蜘蛛毒液中的成分已被證實具有潛在的藥用價值,如鎮痛、抗炎、甚至抗腫瘤等作用。同時,了解蜘蛛毒液的作用機制也為開發新型、環保的生物農藥提供了思路。
總的來說,蜘蛛毒液在捕獵過程中的應用展現了自然界的精妙設計。通過長期的演化,蜘蛛形成了這一高效、多功能的生物武器,不僅確保了它們在生態系統中的生存,也為人類提供了研究和利用的寶貴資源。隨著科技的進步,我們對蜘蛛毒液的了解必將不斷深入,這些知識將為生物學、醫學、農業等多個領域帶來新的發展機遇。
9.4 台灣蜘蛛的主要獵物
台灣蜘蛛的主要獵物種類豐富多樣,反映了島上獨特的生態環境和蜘蛛群落的多樣性。由於台灣地理位置特殊,跨越亞熱帶和熱帶氣候區,加上島嶼地形的多樣性,為各種蜘蛛提供了廣泛的棲息地和豐富的獵物資源。
在台灣的森林生態系統中,許多蜘蛛以各種昆蟲為主要獵物。例如,常見的人面蜘蛛(Nephila pilipes)主要捕食飛蛾、蝴蝶、蜜蜂和其他飛行昆蟲。這種大型蜘蛛結網範圍廣闊,能夠捕獲體型較大的飛行昆蟲。牠們的蛛網具有極強的黏性和韌性,能夠有效地捕獲快速飛行的獵物。
在台灣的低海拔地區和都市環境中,常見的白額高腳蛛(Heteropoda venatoria)則採取不同的捕獵策略。這種蜘蛛不織網,而是主動獵捕各種地面昆蟲,如蟑螂、蟻類和小型甲蟲。牠們憑藉敏捷的身手和靈敏的感官,能夠在夜間有效地捕獲這些活動的昆蟲。
台灣的農業生態系統中,蜘蛛扮演著重要的害蟲控制者角色。例如,縱條銀鱗蛛(Leucauge magnifica)和尖尾銀鱗蛛(Leucauge decorata)常見於稻田和果園中,主要捕食各種農業害蟲,如飛蝨、葉蟬和小型飛蛾。這些蜘蛛通過織造精巧的圓形網來捕獲飛行昆蟲,對維持農業生態平衡起著重要作用。
在台灣的溪流和濕地環境中,一些特化的蜘蛛種類以水生昆蟲為主要獵物。例如,某些漁蛛(Dolomedes)屬的蜘蛛能夠在水面上行走,捕捉水面昆蟲如水黽,有時甚至能捕獲小型魚類和蝌蚪。這些蜘蛛展現了對水生環境的獨特適應。
台灣的山區環境中,如阿里山和玉山地區,一些高海拔適應的蜘蛛種類則以當地特有的昆蟲為食。這些蜘蛛可能捕食高山蝶類、甲蟲和其他適應高海拔環境的昆蟲。這種專門化的捕食行為反映了台灣生態系統的多樣性和複雜性。
值得注意的是,台灣的一些特化蜘蛛種類展現了非常特殊的獵物偏好。例如,臺灣圓光白階蛛(Pasilobus bucculentus)主要以蛾類為食,特別是小型蛾類。這種蜘蛛通過模仿植物構造的特殊網狀結構來吸引和捕獲夜間飛行的蛾類。
在台灣的海岸和紅樹林地區,一些蜘蛛種類適應了鹽分較高的環境,以當地特有的昆蟲和小型甲殼類動物為食。這些蜘蛛的存在強調了台灣生態系統的多樣性,以及蜘蛛在各種環境中的適應能力。
台灣的洞穴環境中,一些特化的蜘蛛種類以洞穴昆蟲為主要獵物。這些蜘蛛可能捕食各種適應黑暗環境的昆蟲,如蟋蟀、蜈蚣和其他節肢動物。洞穴蜘蛛的捕食行為展示了牠們對特殊環境的適應能力。
某些台灣蜘蛛種類還表現出獨特的專門化捕食行為。例如,有些跳蛛(Salticidae)專門捕食蟻類,不僅模仿蟻類的外形和行為,還能夠滲入蟻群中捕食。這種高度專門化的捕食策略反映了蜘蛛與特定獵物之間複雜的共同演化關係。
台灣的一些蜘蛛種類還以其他蜘蛛為食,展現了食同類(cannibalism)和捕食同類(araneophagy)的行為。例如,某些盜網蛛(Theridiidae)科的蜘蛛專門獵捕其他蜘蛛物種,這種行為不僅提供了營養,還可能有助於減少競爭。
在台灣的都市環境中,適應人類住所的蜘蛛種類如臺灣家蛛(Parasteatoda tepidariorum)主要捕食家居常見的小型昆蟲,如蚊子、果蠅和小蛾類。這些蜘蛛在人類環境中起著重要的生態控制作用。
某些台灣蜘蛛種類還表現出季節性的獵物偏好變化。例如,在春季和夏季,當飛行昆蟲數量增加時,織網蜘蛛可能更多地捕食飛蛾和蝴蝶,而在秋冬季節,則可能轉向捕食更多地面活動的昆蟲。
總體而言,台灣蜘蛛的主要獵物反映了島上豐富的生物多樣性和複雜的生態關係。從海岸到高山,從原始森林到都市環境,蜘蛛通過多樣化的捕獵策略和獵物選擇,在各種生態系統中扮演著重要的生態角色。這種多樣性不僅體現了蜘蛛的適應能力,也凸顯了台灣生態系統的獨特性和重要性。深入了解台灣蜘蛛的捕食行為和獵物選擇,對於理解和保護台灣的生態系統具有重要意義。
10. 蜘蛛的種間關係
蜘蛛的種間關係是生態系統中複雜互動網絡的重要組成部分。這些關係涵蓋了多個層面,包括蜘蛛作為捕食者、被捕食者、競爭者以及共生關係的參與者。
作為捕食者,蜘蛛通常處於食物鏈的中間位置,主要捕食各種昆蟲和其他小型節肢動物。然而,蜘蛛本身也是許多動物的獵物,如鳥類、蜥蜴、青蛙和某些哺乳動物。這種雙重身份使蜘蛛在生態系統中扮演著關鍵的平衡角色。
蜘蛛之間的競爭關係也十分普遍,特別是在資源有限的環境中。不同種類的蜘蛛可能為爭奪相同的獵物或棲息地而競爭。例如,在森林環境中,不同種類的織網蛛可能競爭有利的織網位置。
一些蜘蛛物種與其他生物形成了獨特的共生關係。例如,某些蟻擬蛛(Myrmarachne)不僅在外形上模仿蟻類,還能在蟻群中生活而不被發現,這種擬態既保護了蜘蛛免受天敵攻擊,也為其提供了獵食蟻類的機會。
此外,蜘蛛還面臨著來自寄生蟲和寄生蜂的威脅。某些蜂類專門將卵產在蜘蛛體內或蛛卵上,幼蟲孵化後以蜘蛛為食。這種關係展示了自然界中捕食者與被捕食者之間微妙的平衡。
蜘蛛與植物之間也存在複雜的關係。許多蜘蛛依賴植物提供棲息地和織網場所,同時通過捕食害蟲來保護植物。這種間接的互利關係在農業生態系統中尤為重要。
10.1 蜘蛛的天敵
蜘蛛雖然是令人生畏的捕食者,但在自然界中也面臨著來自各種天敵的威脅。這些天敵涵蓋了多個生物類群,從鳥類到昆蟲,甚至包括其他蜘蛛。了解蜘蛛的天敵不僅有助於我們理解生態系統中的複雜關係,還能揭示蜘蛛在演化過程中發展出的各種防禦機制。
鳥類是蜘蛛最主要的天敵之一。許多鳥種將蜘蛛作為重要的食物來源,特別是在繁殖季節,蜘蛛富含蛋白質的身體為鳥類提供了寶貴的營養。例如,山雀科(Paridae)的鳥類經常在樹皮縫隙和樹葉間搜尋蜘蛛。蜂鳥(Trochilidae)雖以花蜜為主食,但也會捕食小型蜘蛛來補充蛋白質。在台灣,常見的綠繡眼(Zosterops japonicus)和紅嘴黑鵯(Hypsipetes leucocephalus)都是熱衷於捕食蜘蛛的鳥類。
爬行動物,尤其是蜥蜴,也是蜘蛛的重要天敵。壁虎(Gekkonidae)憑藉其敏捷的身手和黏性腳掌,能夠在牆壁和天花板上捕捉蜘蛛。在台灣,常見的蝎虎(Hemidactylus frenatus)就經常在人類居住環境中獵食蜘蛛。一些較大型的蜥蜴,如石龍子(Scincidae),也會主動搜尋地面或低矮植被中的蜘蛛。
兩棲動物中,青蛙和蟾蜍是蜘蛛的主要天敵。這些捕食者通常在夜間活動,恰好與許多蜘蛛的活動時間重疊。樹蛙(Hylidae)family的成員尤其善於捕捉棲息在植物上的蜘蛛。在台灣,常見的澤蛙(Fejervarya limnocharis)和中國樹蛙(Hyla chinensis)都會將蜘蛛列入其食譜中。
某些哺乳動物也會捕食蜘蛛,尤其是小型哺乳動物。例如,刺蝟、食蟻獸和某些靈長類動物都會將蜘蛛作為食物來源。在台灣,白鼻心(Paguma larvata)這種果子狸偶爾也會捕食較大型的蜘蛛。蝙蝠雖然主要捕食飛行昆蟲,但一些種類也會捕捉織網的蜘蛛。
在昆蟲世界中,蜘蛛同樣面臨著多種天敵的威脅。泥蜂(Sphecidae)family的成員以捕獵蜘蛛聞名,牠們會將麻痺的蜘蛛帶回巢中作為幼蟲的食物。在台灣,常見的蒼蠅虎(Sceliphron spirifex)就專門獵捕蜘蛛。螳螂也是蜘蛛的天敵,憑藉其強大的前肢和敏捷的身手,能夠輕易地捕獲蜘蛛。
值得注意的是,某些蜂類不僅捕食成年蜘蛛,還會將卵產在蜘蛛體內或蛛卵上。這些寄生蜂的幼蟲會逐漸消耗宿主蜘蛛的身體組織,最終導致蜘蛛死亡。例如,蛛蜂(Pompilidae)family的成員專門獵捕蜘蛛作為其幼蟲的食物來源。
有趣的是,蜘蛛也面臨著來自同類的威脅。某些蜘蛛種類會捕食其他蜘蛛,這種行為稱為捕食同類(araneophagy)。例如,盜網蛛(Theridiidae)family中的某些種類專門獵捕其他蜘蛛。甚至在同一種蜘蛛中,大型個體也可能捕食小型個體,這種現象在蜘蛛的繁殖過程中尤為常見,被稱為性食同類(sexual cannibalism)。
面對如此多樣的天敵,蜘蛛演化出了各種防禦策略。偽裝是一種常見的防禦機制,許多蜘蛛能夠通過模仿樹皮、樹葉或花朵來避免被天敵發現。例如,台灣常見的臺灣圓光白階蛛(Pasilobus bucculentus)就能夠完美地融入樹皮環境中。
一些蜘蛛物種則採取主動防禦策略。例如,某些跳蛛(Salticidae)在面對威脅時會做出威嚇姿態,舉起前足顯示色彩鮮艷的體側。還有一些蜘蛛會在受到驚擾時快速掉落或彈跳,以逃避天敵的捕捉。
某些蜘蛛還發展出了集群生活的策略來增加生存機會。例如,社會性蜘蛛(social spiders)通過共同築巢和協作捕獵來提高對天敵的防禦能力。雖然在台灣不常見,但在其他熱帶和亞熱帶地區,這種社會性蜘蛛的存在為我們提供了了解蜘蛛社會行為演化的寶貴機會。
蜘蛛與其天敵之間的關係不僅體現了自然界的生存競爭,也展示了生態系統中的平衡和適應。這種複雜的互動關係對維持生態系統的穩定性至關重要。通過研究蜘蛛的天敵,我們不僅能夠更好地理解蜘蛛的生態地位,還能深入探討生物多樣性的意義和生態系統的運作機制。在台灣這樣一個生物多樣性豐富的島嶼上,蜘蛛及其天敵之間的關係更是體現了當地生態系統的複雜性和獨特性。
10.2 蜘蛛之間的生存競爭
蜘蛛之間的生存競爭是一個複雜而引人入勝的生態現象,反映了自然界中資源分配和適應性進化的多樣性。這種競爭不僅存在於不同種類的蜘蛛之間,同種蜘蛛個體間也存在激烈的競爭關係。競爭的核心通常圍繞著獵物資源、棲息地和繁殖機會展開,而這些因素又與蜘蛛的生存和繁衍密切相關。
在生態位相似的蜘蛛種類之間,競爭尤為激烈。例如,在台灣的森林環境中,不同種類的織網蛛常常爭奪有利的織網位置。這些位置通常是昆蟲飛行頻繁的區域,如林間空地或溪流上方。人面蜘蛛(Nephila pilipes)和臺灣圓蛛(Argiope aetherea)就經常在這樣的環境中競爭。較大型的金線蛛往往佔據有利位置,而體型較小的圓蛛則被迫選擇次優的位置或尋找其他棲息地。
棲息地的競爭不僅限於織網蛛。在地面活動的獵蛛,如白額高腳蛛(Heteropoda venatoria)和塔蘭托狼蛛(Lycosa tarantula),也會為了優質的獵場而競爭。這些獵場可能是落葉豐富的森林地面,或者是昆蟲活動頻繁的草叢邊緣。擁有更好獵場的蜘蛛往往能夠獲得更多的食物資源,從而在繁殖和生存上佔據優勢。
食物資源的競爭是蜘蛛之間最直接也最激烈的競爭形式之一。在資源有限的環境中,不同種類的蜘蛛可能爭奪相同的獵物。例如,在台灣的農田生態系統中,縱條銀鱗蛛(Leucauge magnifica)和尖尾銀鱗蛛(Leucauge decorata)常常共存,它們都以小型飛蟲為食。當獵物稀少時,這兩種蜘蛛之間的競爭就會加劇。較為強勢的個體可能會搶奪其他蜘蛛已經捕獲的獵物,這種行為在蜘蛛世界中並不罕見。
繁殖機會的競爭在蜘蛛世界中尤為激烈,特別是在雄性蜘蛛之間。許多蜘蛛物種的雄性個體會為了爭奪交配權而進行激烈的競爭。這種競爭可能表現為直接的身體衝突,或者是複雜的求偶展示。例如,臺灣跳蛛(Cosmophasis umbratica)的雄性個體會通過炫耀性的舞蹈和色彩展示來吸引雌性,同時也是在與其他雄性競爭。在這個過程中,體型較大、色彩更鮮艷的雄性往往更容易獲得交配機會。
有趣的是,某些蜘蛛物種還會展現出領地行為,這進一步加劇了競爭。例如,某些長疣蛛(Gasteracantha)種類的雌性會積極防衛自己的網域,驅逐任何入侵者,包括同種的其他個體。這種行為不僅保護了自己的獵場,也減少了潛在的競爭壓力。
在蜘蛛的生活史中,競爭壓力可能因年齡和季節而異。年輕的蜘蛛個體通常面臨更大的競爭壓力,因為它們需要在有限的資源環境中迅速成長。在繁殖季節,競爭往往達到高峰,因為這是決定基因傳遞的關鍵時期。
值得注意的是,蜘蛛之間的競爭並不總是直接和對抗性的。許多蜘蛛物種通過生態位分化來減少競爭。例如,在同一棵樹上,不同種類的蜘蛛可能選擇不同的高度或方向來織網,從而減少直接競爭。姬蛛(Theridion fatimae)常常選擇在樹冠層織網,而臺灣圓蛛則偏好較低的位置,這種空間分配有效降低了種間競爭。
某些蜘蛛物種還發展出了特殊的策略來應對競爭。例如,一些小型蜘蛛種類會選擇在大型蜘蛛的網中或網緣生活,這種被稱為蛛網共生(kleptoparasitism)的行為使它們能夠分享大型蜘蛛捕獲的獵物,同時又避免了直接競爭。在台灣,可以觀察到一些細蛛(Argyrodes)種類在人面蜘蛛的大型蛛網邊緣生活的現象。
蜘蛛之間的競爭也可能導致一些有趣的行為適應。例如,某些蜘蛛物種會採取欺騙性的策略來獲取資源。有些雄性蜘蛛會模仿雌性或獵物的行為來接近其他蜘蛛的網,以獲得交配機會或食物。這種策略雖然風險較高,但在資源稀缺的環境中可能是一種有效的生存策略。
在極端情況下,蜘蛛之間的競爭可能導致同類相食。這種行為在許多蜘蛛物種中都有觀察到,特別是在食物稀缺或空間有限的情況下。例如,在實驗室條件下,研究人員觀察到臺灣狼蛛的幼體在高密度環境中會出現同類相食的行為。
蜘蛛之間的競爭不僅影響個體的生存和繁殖,還在更大的尺度上塑造了蜘蛛群落的結構和動態。通過競爭,不同的蜘蛛物種找到了各自的生態位,形成了複雜而平衡的生態系統。這種競爭也是推動蜘蛛演化的重要力量,促使它們不斷發展新的適應性特徵和行為策略。
在台灣這樣一個生物多樣性豐富的島嶼上,蜘蛛之間的競爭關係尤為複雜和多樣。從低地到高山,從原始森林到都市環境,不同的蜘蛛物種在各自的生態位中競爭和共存,構成了台灣獨特的蜘蛛生態景觀。研究和理解這些競爭關係,不僅有助於我們更好地認識蜘蛛的生態學和行為學,也為保護生物多樣性和維護生態平衡提供了重要的科學基礎。
10.3 蜘蛛的寄生蟲和寄生蜂
蜘蛛作為生態系統中的重要捕食者,同時也面臨著來自寄生蟲和寄生蜂的威脅。這些微小但強大的天敵對蜘蛛的生存和繁衍產生深遠影響,形成了複雜而有趣的生態關係。
寄生蟲是一類以蜘蛛為宿主,長期生活在蜘蛛體內或體表的生物。它們從蜘蛛身上獲取營養,同時可能影響蜘蛛的行為和生理功能。常見的蜘蛛寄生蟲包括線蟲、絛蟲和某些真菌。這些寄生蟲通常不會立即致死宿主,而是長期影響蜘蛛的健康和繁殖能力。
在台灣,一種常見的蜘蛛寄生蟲是蜘蛛蠕形蟲(Mermithidae family)。這種線蟲幼蟲寄生在蜘蛛的腹部,隨著蜘蛛的生長而發育。當線蟲成熟時,會破體而出,導致宿主蜘蛛死亡。這種寄生關係對蜘蛛種群的數量和分佈產生顯著影響,特別是在潮濕的環境中。
另一類值得關注的蜘蛛寄生者是真菌。例如,蟲草屬(Cordyceps)的某些種類專門感染蜘蛛。這些真菌的孢子附著在蜘蛛體表,發芽後侵入蜘蛛體內,最終導致宿主死亡。在死亡過程中,真菌會改變蜘蛛的行為,使其爬到植物的高處,以便真菌的子實體能夠更好地釋放孢子。這種行為操縱展示了寄生者與宿主之間的複雜互動。
相較於寄生蟲,寄生蜂對蜘蛛的威脅更為直接和致命。寄生蜂是一類將卵產在蜘蛛體內或蛛卵上的昆蟲,其幼蟲以蜘蛛的組織為食,最終導致宿主死亡。這種關係被稱為寄生擬寄生(parasitoidism),區別於一般的寄生關係。
在台灣,蛛蜂科(Pompilidae)的成員是最著名的蜘蛛寄生蜂。這些蜂類專門獵捕蜘蛛作為其幼蟲的食物來源。例如,台灣常見的藍翅蛛蜂(Cryptocheilus formosicola)會主動搜尋地面活動的蜘蛛,如狼蛛或跳蛛。當找到合適的獵物時,蛛蜂會用螫針刺傷蜘蛛,注入麻痺毒素。隨後,蛛蜂將麻痺的蜘蛛拖到預先準備的巢穴中,並在蜘蛛身上產卵。孵化的蛛蜂幼蟲會逐漸消耗仍然活著但無法動彈的蜘蛛,直到蜘蛛最終死亡。
除了蛛蜂,泥蜂科(Sphecidae)的某些成員也是蜘蛛的重要天敵。在台灣,常見的蒼蠅虎(Sceliphron spirifex)就專門獵捕各種蜘蛛。這種蜂類會建造泥巢,並將麻痺的蜘蛛儲存在巢室中作為幼蟲的食物。一個巢室可能儲存多隻蜘蛛,為發育中的蜂蛹提供充足的營養。
值得注意的是,一些極小型的寄生蜂專門攻擊蜘蛛的卵。例如,屬於卵蜂科(Scelionidae)的某些種類會將卵產在蜘蛛的卵囊內。這些微小的寄生蜂幼蟲以蜘蛛卵為食,從而大大降低了蜘蛛的繁殖成功率。在台灣的農業生態系統中,這種寄生關係對控制某些蜘蛛種群的數量起到了重要作用。
面對這些寄生者的威脅,蜘蛛演化出了多種防禦策略。一些蜘蛛物種會通過頻繁更換網址來減少被寄生蜂發現的機會。另一些則會在產卵時採取特殊的保護措施,如將卵囊隱藏在複雜的結構中,或者持續守衛卵囊直到幼蛛孵化。
有趣的是,某些蜘蛛物種還發展出了與寄生者共存的策略。例如,有些跳蛛(Salticidae)會容忍特定的寄生蟲存在,因為這些寄生蟲可能會分泌某些物質,有助於蜘蛛抵抗其他更危險的病原體。這種複雜的關係展示了自然界中生物間相互作用的多樣性和精妙性。
在台灣的生態系統中,蜘蛛與其寄生者之間的關係對維持生態平衡起著重要作用。這些寄生者通過控制蜘蛛的數量,間接影響了昆蟲群落的結構。同時,寄生者的存在也為其他生物提供了食物來源,如某些鳥類會專門捕食被真菌感染的蜘蛛。
研究蜘蛛的寄生蟲和寄生蜂不僅有助於我們理解生態系統的複雜性,還為生物防治提供了新的思路。例如,了解特定寄生蜂對農業害蟲的控制作用,可以幫助我們開發更環保、更有效的害蟲管理策略。
此外,蜘蛛和寄生者之間的關係也為進化生物學提供了豐富的研究素材。例如,寄生蜂如何精確定位和識別宿主蜘蛛,以及蜘蛛如何進化出應對寄生威脅的機制,這些都是極具吸引力的研究題目。
在台灣這樣一個生物多樣性豐富的島嶼上,蜘蛛與其寄生者之間的關係可能比我們目前所知的更加複雜和多樣。隨著研究的深入,我們或許能夠發現更多獨特的寄生關係和適應機制,這不僅能豐富我們對台灣生態系統的理解,也可能為生物防治和環境保護提供新的思路和方法。
10.4 蜘蛛與植物的關係
蜘蛛與植物之間存在著複雜而有趣的關係,這種關係遠比人們通常認知的要深入和多樣。雖然蜘蛛不直接以植物為食,但植物為蜘蛛提供了棲息地、獵場和保護,而蜘蛛則通過捕食害蟲來保護植物,形成了一種互利共生的關係。
在台灣豐富多樣的生態系統中,蜘蛛與植物的關係表現得尤為明顯。許多蜘蛛物種選擇特定的植物作為其主要棲息地。例如,人面蜘蛛(Nephila pilipes)常常在森林邊緣或開闊地帶的高大植物之間築網。這些植物不僅提供了理想的網址位置,還為金線蛛創造了有利的獵捕環境,因為許多昆蟲會在這些區域活動。
某些蜘蛛物種甚至與特定植物形成了緊密的關聯。在台灣的低海拔山區,可以觀察到臺灣圓光白階蛛(Pasilobus bucculentus)經常選擇特定種類的樹木作為棲息地。這種蜘蛛的體色和形態與樹皮極為相似,展現了高度的擬態適應。這種關係不僅為蜘蛛提供了保護,也使得蜘蛛能夠更有效地捕捉在樹幹上活動的昆蟲。
植物的構造特徵也對蜘蛛的棲息和捕獵策略產生重要影響。例如,台灣常見的縱條銀鱗蛛(Leucauge magnifica)偏好在具有寬大葉片的植物之間築網。這些葉片不僅為蛛網提供了良好的支撐點,還能在風雨中為蛛網提供一定的保護。同時,這種環境也吸引了許多飛行昆蟲,為蜘蛛提供了豐富的食物來源。
在農業生態系統中,蜘蛛與植物的關係更顯重要。許多蜘蛛物種在農作物中扮演著天敵的角色,有效控制害蟲數量。例如,在台灣的水稻田中,塔蘭托狼蛛(Lycosa tarantula)和日本長腳蛛(Tetragnatha maxillosa)等蜘蛛物種能夠捕食多種水稻害蟲,如飛蝨和葉蟬。這種生物防治作用不僅減少了農藥的使用,還維護了農田生態系統的平衡。
某些蜘蛛物種還展現出了對特定植物結構的適應性。例如,臺灣長疣蛛(Gasteracantha kuhlii)常常選擇在具有堅韌葉片的植物上築網。這種蜘蛛的網具有很強的韌性,能夠承受較大的衝擊,而堅韌的葉片為其提供了理想的網絲附著點。這種適應性選擇反映了蜘蛛與植物之間長期共同演化的結果。
有趣的是,一些蜘蛛物種還利用植物的特殊結構來增強其捕獵效率。例如,在台灣的蘭花園中,可以觀察到某些跳蛛(Salticidae)物種專門在蘭花的花瓣上捕獵。這些蜘蛛利用花瓣的顏色和形狀作為掩護,等待前來授粉的昆蟲。這種行為不僅提高了蜘蛛的捕獵成功率,還可能間接影響植物的授粉過程。
植物還為蜘蛛的繁殖提供了重要支持。許多蜘蛛物種選擇在植物的特定部位產卵和養育幼蛛。例如,姬蛛(Theridion fatimae)常常將其卵囊附著在樹葉背面,利用葉片提供的隱蔽和適宜的微環境條件。一些織網蛛則會在植物枝條之間建造特殊的育幼網,為幼蛛提供初期的保護和獵場。
在台灣的紅樹林生態系統中,蜘蛛與植物的關係呈現出獨特的面貌。紅樹林的複雜根系和週期性的潮汐環境為特化的蜘蛛物種提供了獨特的棲息地。例如,某些蟹蛛(Thomisidae)物種專門在紅樹林的葉片上捕獵,它們的體色和形態與紅樹葉片極為相似,展現了高度的環境適應性。
植物的化學防禦也對蜘蛛產生影響。某些植物分泌的化學物質可能吸引或驅離特定的蜘蛛物種。例如,有研究發現,某些散發特殊氣味的植物可能吸引特定的蜘蛛物種在其周圍築網,這可能是因為這些氣味也吸引了大量的昆蟲。這種間接的相互作用展示了生態系統中複雜的化學通訊網絡。
在台灣的高山生態系統中,蜘蛛與植物的關係更顯獨特。高山環境的極端條件促使蜘蛛與植物形成了緊密的依存關係。例如,在玉山地區,某些蜘蛛物種專門棲息在墊狀植物的緊密結構中,這些植物不僅為蜘蛛提供了抵禦強風和低溫的庇護所,還為其創造了獨特的微型生態系統。
蜘蛛還可能在植物的授粉和種子傳播過程中扮演間接角色。雖然蜘蛛本身不直接參與這些過程,但它們的存在可能影響授粉昆蟲和種子傳播者的行為。例如,某些花上的蜘蛛可能捕食部分授粉昆蟲,從而影響植物的繁殖成功率。這種複雜的相互作用凸顯了生態系統中各個組成部分之間的緊密聯繫。
值得注意的是,植物與蜘蛛的關係並非總是正面的。某些植物可能通過物理或化學方式阻礙蜘蛛的活動。例如,具有黏性分泌物的植物可能會困住小型蜘蛛,而某些植物的茂密葉片可能妨礙蜘蛛築網。這種相互制約的關係反映了自然界中生物間微妙的平衡。
在台灣的城市環境中,蜘蛛與植物的關係也呈現出新的面貌。都市綠化為蜘蛛提供了新的棲息環境,例如公園和行道樹。這些人工栽培的植物為城市蜘蛛群落的形成和維持提供了重要支持,同時蜘蛛也在控制城市害蟲方面發揮了作用。
蜘蛛與植物的關係不僅體現了生態系統的複雜性,也為生態農業和城市綠化提供了重要啟示。通過理解和利用這種關係,我們可以更好地設計農業生態系統和城市綠地,從而促進生物多樣性保護和生態平衡的維護。在台灣這樣一個生物多樣性豐富的島嶼上,深入研究蜘蛛與植物的關係不僅有助於我們更好地理解本地生態系統,還可能為全球生態研究提供寶貴的案例和見解。
10.5 台灣蜘蛛的共生關係
台灣蜘蛛的共生關係是一個極為有趣且複雜的生態學主題。在台灣豐富多樣的生態環境中,蜘蛛與其他生物之間形成了許多獨特的共生關係,這些關係不僅反映了蜘蛛的適應能力,也展現了生態系統的複雜性。
其中一個典型的例子是某些蜘蛛與螞蟻之間的共生關係。在台灣的熱帶和亞熱帶森林中,有一些蜘蛛物種專門棲息在螞蟻巢穴附近或甚至直接生活在螞蟻巢中。這些蜘蛛通過模仿螞蟻的外形和行為來避免被發現,同時它們可以捕食進入螞蟻巢穴的小型昆蟲,或者偷食螞蟻儲存的食物。這種關係被稱為擬寄生共生,蜘蛛從中獲益,而對螞蟻的影響則相對較小。
另一個值得注意的共生關係是某些蜘蛛與植物之間的互動。台灣的一些蘭花物種,如蝴蝶蘭,其花朵結構複雜,常常成為小型昆蟲的棲息地。一些體型較小的蜘蛛物種會選擇在這些花朵中築巢,它們不僅可以獲得良好的隱蔽環境,還能捕食前來訪花的昆蟲。這種關係對植物來說也是有利的,因為蜘蛛可以幫助控制可能危害花朵的害蟲數量。
在台灣的沿海地區,一些蜘蛛物種與紅樹林形成了緊密的共生關係。這些蜘蛛適應了潮汐環境,能夠在紅樹林的樹幹和氣根上築網。當潮水上漲時,它們會迅速移動到較高的位置,等潮水退去後再回到原來的位置。這種適應不僅展現了蜘蛛的生存智慧,也為紅樹林生態系統增添了一個重要的捕食者角色。
台灣的一些蜘蛛物種還與某些鳥類形成了有趣的共生關係。例如,有些織網蜘蛛會選擇在鳥巢附近築網,利用鳥類活動吸引來的飛蟲作為食物來源。同時,這些蜘蛛的存在也為鳥巢提供了一定程度的保護,因為它們可以捕食一些可能危害鳥蛋或幼鳥的小型昆蟲。
在台灣的農業生態系統中,蜘蛛與農作物之間也存在著一種特殊的共生關係。許多農民認識到蜘蛛在害蟲控制中的重要作用,因此會有意識地保護田間的蜘蛛族群。這些蜘蛛通過捕食如蚜蟲、粉蝨等農業害蟲,幫助減少農藥的使用,從而形成了一種人為促成的共生關係。
此外,台灣的一些洞穴蜘蛛與蝙蝠之間也形成了有趣的共生關係。這些蜘蛛常常在洞穴中築網,捕食被蝙蝠驚擾而飛起的昆蟲,或者捕食落在地上的蝙蝠糞便中孵化出的昆蟲幼蟲。這種關係為洞穴生態系統的能量流動提供了一個重要環節。
台灣蜘蛛的共生關係還包括與微生物的互動。最近的研究發現,某些蜘蛛體內存在著特定的共生細菌,這些細菌可能在蜘蛛的消化、免疫甚至是蛛絲生產過程中扮演重要角色。這種微觀層面的共生關係揭示了蜘蛛生理適應的複雜性,也為未來的生物技術研究提供了新的方向。
總的來說,台灣蜘蛛的共生關係反映了自然界的相互依存和平衡。這些關係不僅豐富了台灣的生物多樣性,也為生態系統的穩定性做出了重要貢獻。深入研究這些共生關係,不僅可以增進我們對蜘蛛生態的理解,也能為生態保護和可持續農業等領域提供寶貴的啟示。
11. 蜘蛛的棲息地生態
蜘蛛的棲息地生態是理解這些八足動物生存策略的關鍵。蜘蛛展現出驚人的適應能力,能夠在各種環境中生存繁衍。從熱帶雨林到乾旱沙漠,從高山峻嶺到深海洞穴,幾乎每個角落都能找到蜘蛛的蹤跡。
蜘蛛選擇棲息地時考慮多個因素,包括食物來源、築網條件、捕食者威脅和繁殖需求等。不同種類的蜘蛛有其特定的棲息地偏好。例如,園蛛科(Araneidae)的成員通常喜歡在開闊地區築網,如草地或灌木叢間;而跳蛛科(Salticidae)的蜘蛛則偏好在樹皮、岩石表面等粗糙平面上活動。
某些蜘蛛種類展現出獨特的棲息地適應。水蜘蛛(Argyroneta aquatica)能在水下生活,通過在水中織造充滿空氣的鐘形網來呼吸。另一方面,沙漠中的某些蜘蛛物種能夠在極端乾燥和高溫環境中生存,它們會挖掘深洞或利用植物遮蔭來調節體溫。
蜘蛛的擴散能力對其棲息地選擇也有重要影響。許多蜘蛛幼體能夠利用絲線進行「氣球飛行」,藉助氣流傳播到遠處。這種行為使得蜘蛛能夠快速佔據新的棲息地,特別是在生態系統受到干擾後的恢復過程中。
人類活動對蜘蛛的棲息地產生了深遠影響。城市化進程創造了新的棲息環境,如建築物外牆和公園綠地,某些適應能力強的蜘蛛物種成功地在這些人造環境中繁衍。然而,棲息地破碎化和污染等問題也威脅著許多蜘蛛種類的生存,特別是那些對特定環境要求較高的物種。
11.1 蜘蛛的棲息地類型
蜘蛛是地球上分布最廣泛的生物之一,它們能夠適應各種不同的環境條件,因此在多樣化的棲息地中都能找到它們的身影。蜘蛛的棲息地類型非常豐富,從熱帶雨林到極地苔原,從海岸線到高山頂峰,甚至包括人類居住的都市環境,都有蜘蛛的存在。
森林是蜘蛛最為常見和多樣化的棲息地之一。在熱帶雨林中,蜘蛛的多樣性達到頂峰。這裡的環境複雜,為不同種類的蜘蛛提供了豐富的生態位。例如,樹冠層中的蜘蛛可能專門捕食飛蟲,而地表的蜘蛛則以小型節肢動物為食。在溫帶落葉林中,蜘蛛的種類雖然不如熱帶雨林豐富,但仍然存在著獨特的適應性。一些蜘蛛會在落葉層中築巢,利用枯葉作為天然的偽裝。
草原和草甸是另一類重要的蜘蛛棲息地。這裡的開闊環境非常適合織網蜘蛛,如園蛛科的成員。它們可以在草莖之間織造大型圓形網,捕捉飛行昆蟲。同時,草叢也為一些地棲蜘蛛提供了良好的隱蔽處。在北美的大草原上,研究人員發現,蜘蛛的多樣性和豐度與草原的植被結構密切相關。
沙漠環境雖然看似惡劣,但也有許多蜘蛛適應了這種極端條件。沙漠蜘蛛通常具有特殊的生理和行為適應,例如能夠長時間忍受缺水,或者在白天高溫時躲在地下。一些沙漠蜘蛛,如澳大利亞的輪蛛(Carparachne aureoflava),能夠通過在沙丘上翻滾來快速逃離天敵。
濕地和水域環境也是許多蜘蛛的家園。一些蜘蛛種類專門適應了這種潮濕的環境。例如,泥灘上常見的狼蛛能夠在水面上奔跑,甚至短暫潛入水中。而前面提到的水蜘蛛(Argyroneta aquatica)則更進一步,幾乎整個生命週期都在水下度過,它們會在水中築造一個充滿空氣的「潛水鐘」作為棲息所。
高山地區雖然環境嚴酷,但仍有一些蜘蛛種類能夠在這裡生存。這些高山蜘蛛通常具有特殊的適應性,如能夠抵抗低溫和強紫外線輻射。有些高山蜘蛛物種甚至能在冰川邊緣生活,它們的體內含有一種類似於防凍劑的物質,使其能夠在接近零度的溫度下活動。
洞穴環境是另一種特殊的蜘蛛棲息地。洞穴蜘蛛通常具有特殊的形態特徵,如退化的眼睛和較長的附肢。這些特徵使它們能夠更好地適應黑暗和狹窄的空間。一些洞穴蜘蛛甚至完全失去了視力,而發展出更敏銳的觸覺和振動感知能力。
海岸線和潮間帶也是一些特殊蜘蛛種類的棲息地。這裡的蜘蛛必須適應潮汐的周期性變化和高鹽度環境。例如,一些海濱蜘蛛能夠在退潮時在岩石縫隙中活動,而在漲潮時則躲入氣泡中呼吸。
人類建造的環境,如城市和農田,也成為了許多蜘蛛的新棲息地。都市蜘蛛通常具有較強的適應能力,能夠在建築物的縫隙、花園和公園中生存。一些研究表明,城市環境中的蜘蛛可能比自然環境中的同類生長得更快、體型更大,這可能是由於城市中食物較為豐富且捕食者較少。
農業生態系統是另一種重要的人為蜘蛛棲息地。在農田中,蜘蛛扮演著重要的生物防治角色。不同的農作物為不同類型的蜘蛛提供了棲息環境。例如,在水稻田中,一些蜘蛛物種能夠在稻株間築網捕捉害蟲,而另一些則在田埂上活動。
地下環境,如土壤和落葉層,也是許多蜘蛛的重要棲息地。這些地棲蜘蛛通常體型較小,色彩暗淡,它們在分解者食物網中扮演著重要角色。一些研究發現,地下蜘蛛的多樣性和豐度可以反映土壤健康狀況。
值得注意的是,許多蜘蛛物種並不局限於單一類型的棲息地,而是能夠適應多種環境。這種適應性使得蜘蛛能夠在環境變化時迅速調整。例如,一些原本生活在森林邊緣的蜘蛛物種能夠成功地適應城市公園環境。
蜘蛛的棲息地選擇還與其生活史階段密切相關。許多蜘蛛在幼體和成體階段會選擇不同的微棲息地。例如,一些織網蜘蛛的幼體可能prefer較低矮的植被,而成體則會在較高處築網。這種棲息地轉換有助於減少同種間的競爭,也使得蜘蛛能夠更好地利用環境資源。
11.2 蜘蛛的棲息地選擇
蜘蛛的棲息地選擇是一個複雜而精密的過程,涉及多個因素的權衡。這個過程不僅反映了蜘蛛的生理需求和行為特性,還展現了它們對環境的敏銳感知和適應能力。蜘蛛在選擇棲息地時,主要考慮食物可用性、築網條件、捕食風險、微氣候、繁殖機會等因素。
食物可用性是蜘蛛選擇棲息地的首要考慮因素之一。不同種類的蜘蛛有不同的獵食策略和食物偏好,這直接影響了它們的棲息地選擇。例如,織網蜘蛛傾向於選擇飛蟲較多的開闊區域,如林間空地或草地邊緣。這些地方不僅有充足的獵物,還提供了良好的築網空間。相比之下,一些地棲蜘蛛如狼蛛科的成員,則偏好在落葉層或低矮植被中活動,那裡有豐富的地面節肢動物可供捕食。
築網條件對於織網蜘蛛來說是棲息地選擇的關鍵因素。不同種類的織網蜘蛛對築網環境有特定的要求。例如,金蛛科的蜘蛛通常會在樹木或灌木之間架設大型的圓形網,因此它們需要選擇有適當間距的植被環境。相反,一些細網蛛科的蜘蛛則專門在岩石表面或樹皮縫隙中織造不規則的網,因此它們會選擇這類表面粗糙的環境。
捕食風險也是蜘蛛考慮的重要因素。蜘蛛不僅是捕食者,也是許多動物的獵物。因此,它們在選擇棲息地時會權衡獵物豐富度和被捕食的風險。一些蜘蛛會選擇有良好隱蔽條件的地方,如植被茂密處或岩石縫隙。例如,跳蛛科的一些成員會選擇樹皮或葉片背面作為棲息地,這不僅提供了良好的隱蔽,還為它們的跳躍捕獵提供了有利的平台。
微氣候條件對蜘蛛的棲息地選擇有重要影響。溫度、濕度、光照等因素都會影響蜘蛛的生理活動和生存狀況。不同種類的蜘蛛對這些因素的耐受範圍和偏好各不相同。例如,一些喜熱的蜘蛛會選擇陽光直射的地方,而另一些則偏好陰涼潮濕的環境。在沙漠環境中,一些蜘蛛會選擇在白天躲在石頭下或洞穴中,以避免極端高溫和乾燥。
繁殖需求也是蜘蛛選擇棲息地的重要考量。許多蜘蛛物種在繁殖季節會特意選擇特定的環境。例如,一些蜘蛛會在產卵前尋找安全的隱蔽處,如樹葉捲曲處或岩石縫隙。有些雄性蜘蛛會選擇特定的位置等待雌性,或者在雌性附近築巢以增加交配機會。
蜘蛛的棲息地選擇還與其生活史階段密切相關。幼蛛和成蛛可能會選擇不同的微棲息地。這種選擇有助於減少同種競爭,並滿足不同生長階段的特定需求。例如,一些織網蜘蛛的幼體可能會選擇較低矮的植被築網,而成體則會在較高處築網。這種垂直分層不僅減少了種內競爭,還使得蜘蛛能夠更有效地利用環境資源。
棲息地的穩定性和可預測性也是蜘蛛考慮的因素。一些蜘蛛傾向於選擇相對穩定的環境,這樣可以減少重新選擇和適應新環境的能量消耗。然而,也有一些蜘蛛物種能夠快速適應變化的環境,它們可能會選擇較不穩定但資源豐富的棲息地。
人類活動對蜘蛛的棲息地選擇產生了重大影響。城市化和農業活動創造了新的棲息環境,一些適應能力強的蜘蛛物種成功地在這些人為環境中繁衍。例如,家蝕蛛科的一些成員已經完全適應了室內環境,它們幾乎只在人類建築物中生活。同時,一些原本生活在自然環境中的蜘蛛也逐漸適應了城市公園和花園等半自然環境。
蜘蛛的分散能力也影響其棲息地選擇。許多蜘蛛物種,特別是幼蛛,能夠通過「氣球飛行」的方式進行長距離分散。這種能力使得蜘蛛能夠快速佔據新的適宜棲息地,特別是在生態系統受到干擾後的恢復過程中。然而,這種分散也帶來了風險,因為蜘蛛可能會落在不適合生存的環境中。
值得注意的是,蜘蛛的棲息地選擇並非一成不變。隨著環境條件的變化,如季節更替或氣候變化,蜘蛛可能會調整其棲息地選擇。例如,一些蜘蛛在冬季會遷移到更溫暖或有更好保護條件的微棲息地。此外,棲息地的選擇還可能受到種間競爭的影響。當多個蜘蛛物種共存時,它們可能會通過選擇不同的微棲息地來減少競爭。
蜘蛛的棲息地選擇能力不僅體現了它們的生態適應性,也反映了生態系統的健康狀況。通過研究蜘蛛的棲息地選擇模式,科學家可以更好地理解生態系統的結構和功能,這對於生態保護和管理具有重要意義。例如,通過監測特定蜘蛛物種的分布變化,可以評估棲息地質量的變化或環境污染的影響。
11.3 台灣蜘蛛的微棲息地選擇
台灣蜘蛛的微棲息地選擇是一個豐富多樣且極具地方特色的研究領域。由於台灣獨特的地理位置和多變的地形,從海岸到高山,從平原到森林,為蜘蛛提供了各種各樣的微棲息地。這些微棲息地的選擇反映了台灣蜘蛛對當地生態環境的精細適應。
在台灣的低海拔地區,如沿海平原和丘陵地帶,許多蜘蛛選擇人為環境作為其微棲息地。例如, 白額高腳蛛(Heteropoda venatoria)常見於建築物的牆角和天花板。這種選擇不僅為它們提供了良好的隱蔽處,還能接近人類活動吸引來的昆蟲。同樣,在都市公園和庭院中,常見台灣金蛛(Nephila pilipes)在樹木間架設大型網狀。這些地方不僅有充足的築網空間,還能捕捉到豐富的飛蟲。
台灣的農業生態系統為許多蜘蛛提供了獨特的微棲息地。在水稻田中,台灣中型狼蛛(Lycosa pseudoannulata)常選擇田埂作為其活動場所。這種微棲息地選擇使得它們既能捕捉水田中的昆蟲,又能避免長期浸水。而在果園和茶園中,如台灣草蛛(Agelena labyrinthica)則偏好在低矮植被間織造漏斗狀網,這種網不僅能有效捕捉地面活動的昆蟲,還能為蜘蛛提供隱蔽和保護。
台灣的森林生態系統為蜘蛛提供了多層次的微棲息地。在森林的樹冠層,人面蜘蛛(Nephila pilipes)常在較高的樹枝間織造大型圓網。這種位置不僅有利於捕捉飛行昆蟲,還能避免地面捕食者的威脅。在林下層,台灣條紋新園蛛(Neoscona adianta)則偏好在灌木叢中築網,這種微棲息地既有足夠的空間織網,又能提供良好的隱蔽。
在台灣的高山生態系統中,蜘蛛的微棲息地選擇展現出特殊的適應性。例如,在玉山園區的高海拔地區,一些耐寒的跳蛛種類如玉山跳蛛(Euophrys yushanensis)選擇岩石表面或高山植物叢中作為其微棲息地。這些地方不僅能提供良好的日照以維持體溫,還能為它們的跳躍捕獵提供有利的平台。
台灣的溪流和濕地環境也為某些蜘蛛提供了獨特的微棲息地。例如,台灣蹼足蛛(Dolomedes raptor)常見於溪流邊緣或靜水區域。這種微棲息地選擇使得它們能夠在水面上行走,捕捉水生昆蟲或小魚。而在沼澤地區,一些網蛛種類如台灣漁網蛛(Tetragnatha maxillosa)則選擇在水生植物間架設網狀,這不僅方便捕捉水面上的昆蟲,還能避免地面捕食者。
台灣的海岸線和潮間帶也是一些特殊蜘蛛的微棲息地。例如,在東北角海岸的岩石地帶,常見一種適應鹽分的狼蛛(Arctosa pulverulenta)。這種蜘蛛選擇在岩石縫隙中築巢,這樣既能避免被海浪沖走,又能在退潮時出來捕食小型甲殼類和昆蟲。
在台灣的洞穴環境中,也有一些特化的蜘蛛種類。例如,在墾丁國家公園的石灰岩洞穴中,發現了一種洞穴適應型的長腳蛛(Pholcus medog)。這種蜘蛛選擇在洞穴深處的岩壁上築網,適應了完全黑暗的環境,主要依靠振動感知來捕捉獵物。
台灣的都市化地區為一些適應性強的蜘蛛提供了新的微棲息地。例如,在台北市的公園和綠地中,常見溫室希蛛(Parasteatoda tepidariorum)在人造構築物如欄杆、路燈等處築網。這種微棲息地選擇不僅為它們提供了穩定的結構支撐,還能利用人工照明吸引來的昆蟲作為食物來源。
值得注意的是,台灣蜘蛛的微棲息地選擇還受到季節變化的影響。例如,在夏季,許多蜘蛛會選擇較為陰涼的微棲息地以避免過熱。而在冬季,一些蜘蛛則會遷移到較為溫暖或有更好保護條件的微環境中。這種季節性的微棲息地轉換展現了台灣蜘蛛對當地氣候的適應能力。
此外,台灣蜘蛛的微棲息地選擇還反映了島嶼生態系統的特殊性。由於台灣的地理隔離,一些特有種蜘蛛發展出了獨特的微棲息地偏好。例如,台灣特有的高砂姬蛛(Theridion takayense)只在特定的高山植被中發現,這種高度專一化的微棲息地選擇可能是長期演化適應的結果。
台灣蜘蛛的微棲息地選擇研究不僅有助於了解當地生態系統的結構和功能,還為生態保育提供了重要參考。通過了解不同蜘蛛種類的微棲息地需求,我們可以更好地保護和管理台灣的自然環境,維護生物多樣性。同時,這些研究也為城市規劃和綠化提供了有價值的建議,如何在都市環境中營造適合本地蜘蛛生存的微棲息地,從而提高城市生態系統的健康度。
11.4 蜘蛛的擴散機制
蜘蛛的擴散機制是一個複雜而有趣的生態過程,它不僅影響蜘蛛的分布範圍,還對整個生態系統的結構和動態產生重要影響。蜘蛛擁有多種擴散策略,這些策略隨著物種、年齡階段和環境條件的不同而有所變化。
最為人所知的蜘蛛擴散機制是「氣球飛行」(Ballooning)。這種方法主要被幼蛛和一些小型成蛛採用。蜘蛛會爬到高處,如草尖或樹枝頂端,然後釋放一條或多條絲線。當氣流足夠強勁時,絲線會帶著蜘蛛升空。這種飛行方式可以讓蜘蛛傳播到很遠的地方,有時甚至可以跨越海洋。例如,在夏威夷群島上發現的一些蜘蛛種類,就是通過這種方式從遠處的大陸飄過來的。氣球飛行雖然有效,但也充滿風險,因為蜘蛛無法控制其最終落地的位置。
除了空中擴散,陸地爬行也是蜘蛛重要的擴散方式,尤其對於較大型的蜘蛛來說。例如,狼蛛科的成員通常通過地面爬行來擴散。這種方式雖然速度較慢,但允許蜘蛛更精確地選擇棲息地。在複雜的地形中,如森林地面或岩石區域,爬行擴散使蜘蛛能夠適應性地選擇微棲息地。
水體也為蜘蛛提供了擴散途徑。一些蜘蛛能夠在水面上行走或漂浮,利用水流進行被動擴散。例如,某些漁蛛科的成員不僅能在水面上行走,還能潛入水中。這種能力使它們能夠順流而下,佔據新的水域棲息地。在洪水期間,許多陸生蜘蛛也會被水流帶到新的區域,這種非自願的擴散在某些情況下可能導致種群的重新分布。
人類活動也成為蜘蛛擴散的重要途徑。隨著全球貿易和旅遊的增加,許多蜘蛛種類被無意中引入到新的地區。例如,香蕉蜘蛛(Phoneutria sp.)就經常隨著水果貨物被運送到世界各地。此外,一些適應都市環境的蜘蛛種類,如家蝕蛛(Tegenaria domestica),也經常隨著人類遷移而擴散。
蜘蛛的擴散行為常常與其生活史階段密切相關。許多種類在幼蛛階段更傾向於進行長距離擴散,而成蛛則較少進行大規模移動。這種策略可以減少種內競爭,並增加基因流動。例如,園蛛科的許多種類,幼蛛會進行氣球飛行,而成蛛則傾向於在有限的區域內活動。
蜘蛛的擴散還受到環境因素的強烈影響。氣象條件,如風向和風速,直接影響氣球飛行的效果。溫度和濕度變化也會刺激某些蜘蛛種類進行擴散。例如,在乾旱季節,一些蜘蛛可能會遷移到更濕潤的區域。此外,棲息地的破碎化和環境變化也可能迫使蜘蛛進行擴散以尋找適宜的新棲息地。
值得注意的是,不同蜘蛛種類的擴散能力差異很大。一些蜘蛛物種具有很強的擴散能力,能夠快速佔據新的棲息地。例如,黃褐歡蛛(Latrodectus geometricus)就因其強大的適應能力和擴散能力,在全球多個地區成為入侵種。相比之下,一些特化程度高的蜘蛛種類,如洞穴蜘蛛,其擴散能力可能非常有限,這使得它們更容易受到棲息地破壞的威脅。
蜘蛛的擴散機制還與其社會行為有關。雖然大多數蜘蛛是獨居的,但有些種類表現出一定程度的社會性。例如,某些社會性蜘蛛種類可能會整群遷移到新的棲息地。這種集體擴散行為不僅增加了生存機會,還能在新環境中快速建立穩定的種群。
蜘蛛的擴散還可能受到其他生物的影響。例如,一些蜘蛛可能會附著在鳥類或大型昆蟲的身上進行被動擴散。雖然這種情況不太常見,但它為一些蜘蛛提供了跨越地理屏障的機會。
蜘蛛的擴散機制對生態系統產生深遠影響。作為重要的捕食者,蜘蛛的擴散可以影響獵物種群的動態和分布。同時,蜘蛛也是許多鳥類和其他動物的食物來源,其擴散模式可能影響這些捕食者的分布。此外,蜘蛛通過擴散可以在新的環境中建立種群,這對維持和恢復生態系統的平衡具有重要作用。
研究蜘蛛的擴散機制不僅有助於理解生態系統的動態,還對保育工作具有重要意義。通過了解蜘蛛的擴散模式,我們可以更好地預測和管理入侵種的擴散,保護瀕危蜘蛛種類,以及在生態修復項目中合理規劃蜘蛛的重新引入。例如,在設計生態廊道時,考慮不同蜘蛛種類的擴散能力和需求,可以提高廊道的有效性。
12. 蜘蛛在生態系統中的作用
蜘蛛在生態系統中扮演著多重而重要的角色,其影響遍及整個生態網絡。作為廣泛分布的捕食者,蜘蛛對維持生態平衡具有關鍵作用。它們主要通過控制昆蟲和其他小型節肢動物的數量來影響生態系統。
在農業生態系統中,蜘蛛是天然的害蟲控制者。例如,在台灣的水稻田中,狼蛛和園蛛能有效捕食害蟲,減少農藥使用。蜘蛛還是食物網中的重要一環,它們既是捕食者,也是其他動物如鳥類和小型哺乳動物的食物來源,促進了能量在不同營養級間的流動。
蜘蛛對生態系統的影響還體現在它們的網對環境的改變。蛛網不僅捕捉昆蟲,還能截留空氣中的花粉和孢子,影響植物的傳粉和繁殖。某些蜘蛛物種,如金蛛,其大型網甚至能改變局部的微氣候條件。
此外,蜘蛛對生態系統的敏感性使它們成為環境變化的良好指示物。例如,某些蜘蛛物種對污染和棲息地破壞特別敏感,其數量變化可反映環境質量。在城市生態系統中,蜘蛛的多樣性常被用作評估綠地健康的指標之一。
12.1 蜘蛛在食物網中的位置
蜘蛛在食物網中占據著獨特而重要的位置,它們是生態系統中不可或缺的一環。作為廣泛分布的捕食者,蜘蛛在多個營養級之間扮演著關鍵的中介角色,影響著整個生態系統的能量流動和物質循環。
在大多數陸地生態系統中,蜘蛛主要作為次級消費者出現。它們捕食各種初級消費者,如草食性昆蟲、小型節肢動物等。例如,在台灣的森林生態系統中,金蛛科的成員如人面蜘蛛(Nephila pilipes)常常在樹木間織造大型網狀,捕捉飛蛾、蝴蝶等飛行昆蟲。這種捕食行為不僅控制了昆蟲的數量,還間接影響了植物的生長狀況。
然而,蜘蛛的食物網位置並非固定不變。某些蜘蛛物種可能在不同情況下扮演不同的角色。例如,一些大型蜘蛛如台灣巨蛛(Araneus ventricosus)有時會捕食小型蜥蜴或幼鳥,此時它們就上升到了更高的營養級,成為三級消費者。這種靈活性使得蜘蛛能夠適應不同的生態環境,並在食物網中發揮多重作用。
蜘蛛在食物網中的位置還與其生活史階段密切相關。幼蛛通常捕食更小的獵物,如蚜蟲或薊馬,而成熟的蜘蛛則能夠捕捉更大的獵物。這種隨年齡變化的食性使得同一種蜘蛛在其生命週期的不同階段可能占據食物網中的不同位置。
值得注意的是,蜘蛛不僅是捕食者,它們自身也是其他動物的食物來源。許多鳥類、蜥蜴、青蛙,甚至一些小型哺乳動物都以蜘蛛為食。例如,在台灣的森林中,綠繡眼(Zosterops japonicus)常常捕食小型蜘蛛作為其蛋白質來源。這種被捕食的角色使得蜘蛛成為連接低營養級和高營養級生物的重要橋樑。
蜘蛛在食物網中的位置還體現了能量在生態系統中的流動。由於蜘蛛的新陳代謝效率相對較高,它們能夠有效地將捕獲的獵物轉化為自身的生物量。這種高效的能量轉換使得蜘蛛成為生態系統中能量流動的重要節點。
在某些特殊的生態系統中,蜘蛛可能占據食物網的頂端位置。例如,在一些遠離大陸的小島嶼生態系統中,由於缺乏大型捕食者,蜘蛛可能成為食物鏈的頂級掠食者。這種情況在台灣的一些離島如蘭嶼或綠島的某些微生態系統中也可能存在。
蜘蛛在食物網中的位置還受到季節變化的影響。在台灣,春季和秋季通常是蜘蛛活動的高峰期,此時它們在食物網中的作用更為顯著。例如,在秋季,當許多昆蟲開始為冬季做準備時,蜘蛛的捕食壓力可能對這些昆蟲種群的冬季存活率產生重要影響。
人類活動也會影響蜘蛛在食物網中的位置。例如,在農業生態系統中,殺蟲劑的使用可能會減少蜘蛛的食物來源,從而改變它們在食物網中的功能。另一方面,有機農業的推廣可能會增加蜘蛛在害蟲控制中的重要性,強化其在農業食物網中的位置。
蜘蛛在食物網中的位置還體現在其對生態系統穩定性的貢獻。作為廣泛的捕食者,蜘蛛能夠同時控制多種獵物的數量,這種多樣化的捕食壓力有助於維持生態系統的平衡。例如,在台灣的低海拔森林中,多種蜘蛛共同存在,它們捕食不同的昆蟲物種,共同維持了昆蟲群落的多樣性。
蜘蛛的食物網位置還與其特殊的捕食策略有關。不同的蜘蛛物種採用不同的捕獵方式,這使得它們能夠利用生態系統中的不同資源。例如,織網蜘蛛如溫室希蛛(Parasteatoda tepidariorum)主要捕捉飛行昆蟲,而地棲蜘蛛如台灣狼蛛(Lycosa coelestis)則主要捕食地面活動的小型節肢動物。這種策略的多樣性使得蜘蛛能夠在食物網的不同部分發揮作用。
蜘蛛在食物網中的位置還反映了生態系統的健康狀況。在健康的生態系統中,蜘蛛種群通常呈現出穩定的數量和多樣性。相反,如果生態系統受到污染或破壞,蜘蛛的數量和多樣性可能會下降,這可能導致食物網結構的變化。因此,蜘蛛常被用作評估生態系統健康的指標之一。
此外,蜘蛛在食物網中的位置還體現了生態系統的連通性。通過捕食各種昆蟲,蜘蛛將不同的生態系統成分連接起來。例如,一隻蜘蛛可能在同一天內捕食來自水生環境的蚊子和陸地環境的螞蟻,從而將這兩個看似分離的生態系統聯繫在一起。
蜘蛛在食物網中的作用還延伸到非營養關係。例如,某些蜘蛛物種的存在可能影響其他動物的行為。在台灣的一些農田中,研究發現蜘蛛的存在會改變某些昆蟲的活動模式,這間接影響了這些昆蟲對作物的危害程度。
最後,值得注意的是,蜘蛛在食物網中的位置並非靜態不變的。隨著環境條件的變化,如氣候變遷或棲息地改變,蜘蛛在食物網中的角色可能會發生調整。這種動態性使得蜘蛛成為研究生態系統應對環境變化的重要對象。
12.2 蜘蛛在害蟲控制中的角色
蜘蛛在害蟲控制中扮演著極其重要且獨特的角色,它們是自然界中最有效的生物防治agents之一。這些八足動物通過其多樣化的捕獵策略和廣泛的食性,在各種生態系統中發揮著控制害蟲數量的關鍵作用。
在農業生態系統中,蜘蛛的害蟲控制作用尤為顯著。以台灣的水稻田為例,多種蜘蛛物種共同構成了一個複雜的天敵系統。台灣中型狼蛛(Lycosa pseudoannulata)是水稻田中常見的地棲蜘蛛,它們主要在田埂和稻田表面活動,捕食如飛蝨、葉蟬等多種水稻害蟲。這種蜘蛛不僅能夠捕食成蟲,還能有效控制這些害蟲的若蟲數量。
另一方面,織網蜘蛛如溫室希蛛(Parasteatoda tepidariorum)則在空中設置天羅地網,捕捉飛行中的害蟲。這些蛛網不僅能夠捕獲如稻飛蝨、稻縱卷葉蟲等飛行害蟲,還能阻礙害蟲在稻田間的移動和擴散。研究表明,在蜘蛛數量較多的稻田中,害蟲的數量明顯低於蜘蛛數量較少的稻田。
蜘蛛在果園和蔬菜園中的害蟲控制作用同樣不容忽視。在台灣的柑橘園中,常見的黑斑園蛛(Araneus mitificus)能夠有效捕食如柑橘木蝨、柑橘潛葉蛾等害蟲。這些蜘蛛通常在樹冠間織網,不僅能捕獲成蟲,還能阻止害蟲在果樹間的擴散。
蜘蛛的害蟲控制效果還體現在其獨特的捕食行為上。許多蜘蛛物種具有過量捕殺(Surplus killing)的行為,即它們捕獲的獵物數量遠超過其實際food需求。這種行為雖然看似浪費,但在害蟲控制中卻發揮了重要作用。例如,在台灣的茶園中,研究人員發現一些跳蛛物種能夠在短時間內捕殺大量的茶小綠葉蟬,即使它們並不會完全吃掉這些獵物。
蜘蛛在害蟲控制中的另一個優勢是其廣泛的食性。大多數蜘蛛是機會主義捕食者,能夠捕食多種不同的昆蟲。這種多樣化的食性使得蜘蛛能夠在害蟲種群爆發前就開始發揮控制作用,並在害蟲數量減少時轉而捕食其他昆蟲,從而維持穩定的種群數量。例如,在台灣的蔬菜園中,園蛛科的成員不僅能夠捕食主要害蟲如蚜蟲和粉蝶幼蟲,還能捕食其他潛在的害蟲,從而預防多種害蟲的爆發。
蜘蛛的害蟲控制作用還體現在其對生態系統的穩定作用上。由於蜘蛛能夠同時控制多種害蟲的數量,它們有助於維持昆蟲群落的多樣性和平衡。這種平衡作用可以預防單一害蟲種類的大規模爆發。例如,在台灣的某些有機農場中,研究者發現保持高度的蜘蛛多樣性可以顯著降低害蟲爆發的風險。
值得注意的是,蜘蛛在害蟲控制中的作用並不僅限於直接捕食。蜘蛛的存在還能產生所謂的「非消耗性效應」。許多害蟲物種能夠感知蜘蛛的存在,並改變其行為以避免被捕食。這種行為改變可能包括減少取食時間、改變活動範圍或增加隱蔽行為。例如,在台灣的某些溫室實驗中,研究人員發現即使蜘蛛被限制無法直接捕食,其存在仍能顯著降低某些害蟲對作物的危害程度。
蜘蛛在都市環境中的害蟲控制作用也不容忽視。在台灣的城市綠地和住宅區,蜘蛛對控制蚊子、蒼蠅等都市害蟲起著重要作用。例如,常見於屋簷下的姬蛛網能有效捕捉登革熱病媒蚊。這種自然的害蟲控制不僅有助於維護城市環境衛生,還可能減少殺蟲劑的使用,從而降低環境污染。
然而,蜘蛛在害蟲控制中的作用也面臨著挑戰。過度使用農藥是最主要的威脅之一。許多農藥不僅殺死害蟲,也會殺死蜘蛛等天敵,從而破壞了自然的害蟲控制系統。為了充分發揮蜘蛛的害蟲控制作用,一些台灣的農民開始採用綜合病蟲害管理(IPM)策略,減少農藥使用,為蜘蛛創造更有利的生存環境。
此外,農業景觀的簡化也可能降低蜘蛛的害蟲控制效果。單一作物種植往往導致蜘蛛多樣性下降,從而減弱了整體的害蟲控制能力。為此,一些研究者建議在農田周圍保留自然植被,或在田間種植覆蓋作物,以提供蜘蛛更多樣化的棲息環境。
最後,公眾對蜘蛛的認知也影響著其在害蟲控制中的作用。許多人對蜘蛛存在恐懼或厭惡,可能會主動清除環境中的蜘蛛。為此,台灣的一些環境教育項目開始致力於提高公眾對蜘蛛生態價值的認識,強調其在害蟲控制中的重要作用。
12.3 蜘蛛作為生物指標
蜘蛛作為生物指標在生態學研究和環境監測中扮演著越來越重要的角色。這些八足動物因其獨特的生態特性和對環境變化的敏感性,成為評估生態系統健康狀況的理想指標生物。蜘蛛作為生物指標的應用範圍廣泛,從自然生態系統到人為干擾環境,都能提供有價值的生態信息。
在自然生態系統中,蜘蛛的多樣性和豐富度常被用來評估棲息地的質量和生態系統的整體健康狀況。例如,在台灣的森林生態系統研究中,科學家們發現蜘蛛群落的組成和多樣性能夠反映森林的演替階段和干擾程度。原始林中通常擁有更多樣化的蜘蛛群落,包括一些對環境要求較高的專性物種。相比之下,受到干擾的次生林或人工林中,蜘蛛的多樣性往往較低,且以適應性強的廣布種為主。
蜘蛛對環境污染的反應使其成為評估生態系統受污染程度的有效指標。某些蜘蛛物種對特定污染物特別敏感,可以通過監測這些物種的存在與否或種群數量變化來推斷環境中污染物的水平。例如,在台灣某些工業區附近的研究中發現,重金屬污染較嚴重的區域,地棲蜘蛛的種類和數量明顯減少,而一些耐受性較強的織網蜘蛛則相對更為常見。
在農業生態系統中,蜘蛛群落的變化可以反映農業實踐對生態環境的影響。在台灣的有機農業和常規農業對比研究中,科學家們發現有機農田通常擁有更高的蜘蛛多樣性和豐富度。這不僅反映了減少農藥使用對生態系統的積極影響,也暗示了農田生態系統的整體健康狀況。例如,在台南市的某些水稻田研究中,研究人員發現採用有機耕作的稻田中,不僅蜘蛛的物種數量更多,其中還包括一些對環境質量要求較高的稀有物種。
蜘蛛還被用作評估棲息地破碎化和連通性的指標。由於不同蜘蛛物種具有不同的擴散能力和棲息地需求,通過研究蜘蛛群落的組成可以推斷景觀的連通性狀況。在台灣的都市化研究中,科學家們發現,都市綠地中的蜘蛛多樣性與綠地的大小和連通性密切相關。較大且相互連接的綠地通常擁有更多樣化的蜘蛛群落,包括一些需要較大活動範圍的物種。
蜘蛛對氣候變化的反應也使其成為研究全球變暖影響的重要指標。在台灣的高山生態系統研究中,科學家們通過長期監測高海拔地區的蜘蛛群落變化,發現某些原本只分布在較低海拔的蜘蛛物種正逐漸向高海拔地區擴散。這種分布範圍的變化可能反映了氣候變暖的趨勢。
在生態修復項目中,蜘蛛群落的恢復過程常被用作評估修復成效的指標。例如,在台灣某些礦區的生態重建項目中,研究人員通過監測蜘蛛群落的變化來評估生態系統的恢復情況。隨著植被的逐漸恢復,蜘蛛的多樣性和豐富度也隨之增加,特別是一些對環境質量要求較高的森林物種的重新出現,被視為生態修復成功的重要標誌。
蜘蛛作為生物指標還體現在其對微棲息地變化的敏感性上。不同的蜘蛛物種對微環境條件如溫度、濕度、光照等有不同的偏好和耐受範圍。因此,通過研究特定微棲息地中的蜘蛛群落組成,可以推斷該環境的微氣候特徵。在台灣的洞穴生態研究中,科學家們發現不同深度和濕度的洞穴區域擁有不同的蜘蛛群落,這些差異反映了洞穴內部環境的細微變化。
蜘蛛在指示生態系統功能方面也發揮著重要作用。作為廣泛的捕食者,蜘蛛群落的變化可以反映整個生態系統的營養結構和能量流動狀況。例如,在台灣的森林生態系統中,研究人員發現蜘蛛的生物量和多樣性與其他節肢動物的豐富度密切相關,這種關係可以用來評估生態系統的整體生產力和穩定性。
然而,使用蜘蛛作為生物指標也面臨一些挑戰。首先,蜘蛛的分類鑑定需要專業知識,這可能限制了其在大規模環境監測中的應用。其次,蜘蛛群落可能受到多種因素的影響,如季節變化、取樣方法等,這就要求研究者在解釋數據時需要格外謹慎。
儘管如此,蜘蛛作為生物指標的優勢仍然顯著。它們分布廣泛、對環境變化敏感、採樣相對容易,且在生態系統中扮演重要角色。這些特點使得蜘蛛成為環境監測和生態評估中不可或缺的工具。
在台灣,隨著環境保護意識的提高和生態研究的深入,蜘蛛作為生物指標的應用正在不斷擴大。從自然保護區的生態監測到都市環境的綠地評估,再到農業生態系統的健康診斷,蜘蛛都在發揮著重要的指示作用。這不僅為生態學研究提供了寶貴的數據,也為環境管理和保護政策的制定提供了科學依據。
12.4 蜘蛛與生態系統服務
蜘蛛在生態系統中扮演著重要的角色,為自然環境提供了多種生態系統服務。這些八足動物不僅是食物網中的關鍵一環,還通過其獨特的行為和生理特性,為生態系統的平衡和健康做出了重要貢獻。
首先,蜘蛛是自然界中最重要的捕食者之一,它們通過控制昆蟲和其他小型無脊椎動物的數量,為生態系統提供了調節服務。在農業生態系統中,蜘蛛的這一角色尤為突出。它們能夠有效地控制多種農業害蟲,如蚜蟲、蛾類幼蟲和其他可能危害作物的昆蟲。這種自然的害蟲控制機制不僅能夠減少農藥的使用,還能夠提高農作物的產量和質量,從而為人類提供重要的經濟效益。
此外,蜘蛛還通過其獨特的絲線生產能力為生態系統提供了支持服務。蛛絲不僅是蜘蛛捕獵和築巢的工具,還在整個生態系統中發揮著多種功能。例如,某些鳥類會利用蛛絲來建造巢穴,增強巢的結構強度。一些小型昆蟲和植物種子則可能利用蛛絲進行空中傳播,這有助於生物多樣性的擴散和基因交流。
蜘蛛還為生態系統提供了重要的養分循環服務。當蜘蛛捕食其他昆蟲時,它們實際上在參與生態系統中的能量和物質流動。蜘蛛的排泄物和死亡後的屍體都會被分解者分解,從而將養分重新釋放到土壤中,促進植物生長和其他生物的繁衍。
在某些特定的生態系統中,蜘蛛還可能提供獨特的文化服務。例如,在一些地區,觀察蜘蛛的行為被視為預測天氣變化的一種方法。某些文化中,蜘蛛被視為智慧和創造力的象徵,這些文化價值觀念有助於促進人們對自然的尊重和保護意識。
蜘蛛在生態系統中的存在還可能影響其他生物的行為和分佈,從而間接地提供生態系統服務。例如,某些昆蟲可能會避開有大量蜘蛛出沒的區域,這種行為可能會影響植物的授粉模式或種子傳播方式,進而影響整個生態系統的結構和功能。
在城市生態系統中,蜘蛛的作用同樣不容忽視。它們能夠適應人類建築環境,在建築物內外捕食蚊子、蒼蠅等可能傳播疾病的昆蟲,從而為城市居民提供了一定程度的健康保護服務。此外,城市中的蜘蛛還可以作為環境健康的指標,因為它們對環境污染和生態變化較為敏感。
蜘蛛還通過其獨特的生理特性為生物技術和材料科學領域提供了創新的靈感和資源。蛛絲因其優異的強度和彈性,已成為研究人員開發新型材料的重要參考。這些研究不僅可能帶來經濟效益,還可能為環境友好型材料的開發提供新的方向,從而間接地為生態系統的可持續發展做出貢獻。
值得注意的是,蜘蛛提供的生態系統服務往往是相互關聯和複雜的。例如,蜘蛛通過控制某些植食性昆蟲的數量,可能會間接影響植物的生長和繁殖,這又可能影響到依賴這些植物的其他生物,形成一個複雜的生態網絡。因此,保護蜘蛛及其棲息地不僅對蜘蛛本身有益,還能夠維持整個生態系統的穩定性和功能性。
12.5 台灣生態系統中的蜘蛛
台灣作為一個生物多樣性豐富的島嶼,其獨特的地理位置和多樣化的棲息地為蜘蛛提供了絕佳的生存環境。從海岸線到高山,從原始森林到都市綠地,蜘蛛在台灣的各種生態系統中扮演著不可或缺的角色。
在台灣的森林生態系統中,蜘蛛是維持生態平衡的重要一環。以台灣著名的霧林帶為例,這裡濕潤的環境和豐富的植被為許多特殊的蜘蛛物種提供了理想的棲息地。其中,人面蜘蛛(Nephila pilipes)是一種廣泛分布於台灣中低海拔森林中的大型蜘蛛。這種蜘蛛不僅以其巨大的體型和金黃色的絲網吸引了遊客和研究者的注意,更重要的是,它在控制森林昆蟲數量方面發揮著關鍵作用。金線蛛的大型網可以捕獲各種飛行昆蟲,包括一些可能對植物造成危害的害蟲,從而幫助維持森林生態系統的平衡。
在台灣的農業生態系統中,蜘蛛同樣扮演著重要角色。台灣的水稻田是許多蜘蛛物種的重要棲息地,如縱條銀鱗蛛(Leucauge magnifica)和尖尾銀鱗蛛(Leucauge decorata)。這些蜘蛛通過捕食稻田中的害蟲,如稻飛蝨和稻螟蛾,有效地減少了農藥的使用需求,促進了農業的可持續發展。台灣農業研究單位已經開始重視蜘蛛在生物防治中的應用,並鼓勵農民採用友善蜘蛛的耕作方式,以充分利用這些天然的害蟲控制者。
台灣的沿海生態系統也有獨特的蜘蛛群落。在紅樹林和鹽沼地區,一些特化的蜘蛛物種已經適應了潮汐變化和高鹽環境。例如,某些狼蛛科的物種能夠在潮間帶活動,捕食小型甲殼類和昆蟲,成為連接海洋和陸地生態系統的重要紐帶。這些蜘蛛不僅參與了沿海地區的能量流動,還為研究生物如何適應極端環境提供了寶貴的案例。
在台灣的高山生態系統中,蜘蛛展現了驚人的適應能力。即使在海拔3000米以上的高山草原和岩石地帶,仍能發現一些特化的蜘蛛物種。這些高山蜘蛛通常體型較小,具有特殊的生理機制來應對低溫和強紫外線輻射。它們的存在不僅豐富了高山生態系統的食物網,還為研究生物對極端環境的適應提供了重要的研究對象。
台灣的都市生態系統中,蜘蛛也發揮著重要作用。在公園、花園和建築物周圍,常見的蜘蛛如白額高腳蛛(Heteropoda venatoria)成為控制蚊蠅等城市害蟲的重要力量。這些適應城市環境的蜘蛛不僅提供了生態系統服務,還成為城市居民接觸自然的重要媒介,有助於提高公眾的生態意識。
台灣特有的島嶼生態系統為一些特有種蜘蛛的演化提供了獨特的舞台。例如,臺灣黃條盲蛛(Haplogaleoides formosensis)是台灣特有種,主要分布在中低海拔的森林地區。這類特有種的存在不僅增加了台灣的生物多樣性,還為研究島嶼生物地理學和演化過程提供了重要的研究素材。
在台灣的洞穴生態系統中,一些特化的蜘蛛物種也引起了研究者的關注。這些洞穴蜘蛛通常具有退化的眼睛和淺色的體色,反映了它們對黑暗環境的適應。它們在洞穴食物網中扮演著重要的捕食者角色,同時也是研究生物適應極端環境的重要模式生物。
台灣的溪流和河岸生態系統中,某些蜘蛛物種展現了獨特的生存策略。例如,臺灣溪畫蛛(Neoscona punctigera)常在溪流附近築網,捕捉水生昆蟲和剛羽化的蜉蝣等。這些蜘蛛成為連接水陸生態系統的重要環節,參與了養分在不同生態系統間的傳遞和循環。
值得一提的是,台灣蜘蛛群落正面臨著人為活動帶來的挑戰。棲息地破壞、農藥過度使用和氣候變化等因素都對蜘蛛的生存造成威脅。然而,這些威脅也促使研究者更加關注蜘蛛的保護和研究。例如,台灣的一些自然保護區和國家公園正在進行蜘蛛多樣性調查和長期監測計劃,以更好地了解和保護這些重要的生態系統成員。
台灣蜘蛛在各種生態系統中的存在和作用,不僅反映了島嶼生態系統的複雜性和脆弱性,也凸顯了保護生物多樣性的重要性。通過持續的研究和保護努力,台灣的蜘蛛將繼續在維持生態平衡、促進科學研究和提高環境意識等方面發揮重要作用,成為台灣豐富生態資源的重要組成部分。
13. 蜘蛛保育與環境議題
蜘蛛保育與環境議題是當前生態學研究的重要課題。蜘蛛作為生態系統中的關鍵物種,其數量和多樣性的變化能夠反映環境健康狀況。然而,人類活動對蜘蛛棲息地的破壞、全球氣候變遷以及過度使用農藥等因素,正嚴重威脅著蜘蛛的生存。
棲息地喪失是蜘蛛面臨的最大威脅之一。隨著城市化進程加快,大量森林和草原被開發,導致許多蜘蛛失去了原有的生存空間。例如,台灣的都市擴張已經使得一些特有種蜘蛛的分布範圍急劇縮小。此外,農業集約化也對蜘蛛造成嚴重影響,大規模單一作物種植和頻繁的農藥使用降低了農田生態系統的多樣性,使得許多依賴複雜環境的蜘蛛物種難以生存。
氣候變遷對蜘蛛的影響也不容忽視。溫度和濕度的變化可能導致某些蜘蛛物種的分布範圍發生改變,甚至造成部分物種滅絕。例如,高山地區的特有蜘蛛物種可能因為氣溫上升而失去適宜的棲息環境。另一方面,氣候變遷也可能改變蜘蛛的行為模式和生活史特徵,進而影響整個生態系統的平衡。
面對這些威脅,制定有效的蜘蛛保育策略變得尤為重要。這包括建立保護區、恢復受損棲息地、減少農藥使用,以及加強公眾教育等措施。同時,持續的科學研究對於了解蜘蛛的生態需求和應對環境變化的能力至關重要,這將為制定更精準的保育策略提供科學依據。
13.1 台灣蜘蛛面臨的威脅
台灣作為一個生物多樣性豐富的島嶼,其蜘蛛群落正面臨著多方面的威脅。這些威脅不僅影響蜘蛛的生存,還可能對整個生態系統造成深遠的影響。
首要的威脅來自棲息地的破壞和喪失。隨著台灣經濟的快速發展,城市化進程加快,大量自然棲息地被轉化為建築用地、道路和其他人工設施。例如,在台北盆地周邊,原本豐富的低海拔森林生態系統已經大幅縮減,導致許多依賴這類環境的蜘蛛物種,如白額高腳蛛(Heteropoda venatoria)的生存空間受到嚴重擠壓。同樣,沿海地區的開發也威脅著一些特化的濱海蜘蛛物種的生存。
農業活動的集約化是另一個重要威脅。台灣的農業生產為了提高效率和產量,大量使用化學農藥和化肥。這些化學物質不僅直接毒害蜘蛛,還會通過食物鏈累積,對蜘蛛及其獵物造成長期影響。例如,在台灣中部的果園和茶園中,過度使用農藥已經導致一些常見的園藝蜘蛛,如縱條銀鱗蛛(Leucauge magnifica)的數量明顯減少。此外,單一作物種植模式也降低了農田生態系統的複雜性,減少了蜘蛛的棲息和覓食機會。
氣候變遷對台灣蜘蛛的影響也日益顯著。台灣位於亞熱帶地區,氣候變遷導致的極端天氣事件,如強烈颱風和長期乾旱,直接威脅著蜘蛛的生存。特別是一些分布範圍狹窄的特有種蜘蛛,如只生活在台灣中高海拔山區的特有種,可能因為氣溫上升而失去適宜的棲息環境。氣候變化還可能改變植被結構,間接影響蜘蛛的食物來源和築網場所。
外來入侵種的引入也對台灣本土蜘蛛構成威脅。隨著全球貿易的增加,一些非本土蜘蛛物種被無意中引入台灣。這些外來種可能與本土種競爭資源,或者帶來新的疾病和寄生蟲。例如,近年來在台灣都市地區發現的褐牆紅蜘蛛(Nesticodes rufipes),就是一種來自熱帶美洲的入侵物種,它可能與本土的屋內蜘蛛競爭生存空間。
光污染是一個容易被忽視但對蜘蛛影響重大的威脅。台灣城市和鄉村地區的夜間照明不斷增加,干擾了許多夜行性蜘蛛的正常活動。例如,一些依賴夜間捕食的跳蛛和狼蛛可能因為強烈的人工照明而改變其捕獵行為,這不僅影響蜘蛛的生存,還可能打亂當地的生態平衡。
水污染和土壤污染也是台灣蜘蛛面臨的重要威脅。工業廢水和農業化學品的流失可能導致水體和土壤中累積大量有毒物質。這些污染物可能通過食物鏈對蜘蛛造成慢性毒害,影響其生長、繁殖和存活率。特別是一些生活在水邊或土壤中的蜘蛛物種,如某些狼蛛科的物種,更容易受到這類污染的影響。
台灣特有的地理環境也為蜘蛛帶來了獨特的威脅。頻繁的地質活動,如地震和山崩,可能突然改變或破壞蜘蛛的棲息地。雖然這些自然災害在某種程度上促進了生態系統的更新,但對於一些分布範圍狹窄的特有種蜘蛛來說,可能造成滅頂之災。
人類對蜘蛛的誤解和恐懼也是一種間接的威脅。儘管大多數蜘蛛對人類無害,甚至有益,但許多人仍然對蜘蛛懷有不理性的恐懼。這種恐懼可能導致人們過度使用殺蟲劑,或者在遇到蜘蛛時選擇直接殺死它們,而不是將它們安全地移到戶外。這種行為不僅直接威脅蜘蛛的生存,還可能破壞室內生態系統的平衡。
此外,全球化和城市化進程帶來的生活方式改變也對台灣蜘蛛造成影響。例如,現代建築的密封性更好,減少了蜘蛛進入室內的機會,這對一些已經適應人類住所的蜘蛛物種造成了挑戰。同時,城市綠地的減少和碎片化也限制了蜘蛛的活動範圍和基因交流。
值得注意的是,這些威脅往往是相互關聯和複合作用的。例如,氣候變遷可能加劇棲息地破壞的影響,而污染可能使蜘蛛更容易受到氣候變化的影響。因此,保護台灣蜘蛛群落需要一個綜合的、多角度的方法。這包括加強棲息地保護、推廣生態友好型農業practices、控制外來種入侵、減少污染、加強公眾教育等多方面的努力。同時,持續的科學研究對於理解這些威脅的具體機制和蜘蛛的應對能力也至關重要,這將為制定更有效的保護策略提供科學依據。
13.2 台灣蜘蛛保育策略
台灣的蜘蛛保育策略是一個多層面、綜合性的方案,旨在應對該島嶼獨特的生態環境和面臨的威脅。這些策略不僅關注蜘蛛本身,還考慮了整個生態系統的健康和平衡。
首先,棲息地保護是台灣蜘蛛保育的核心策略之一。台灣已經建立了一系列的國家公園和保護區,如玉山國家公園、墾丁國家公園等,這些區域為許多蜘蛛物種提供了安全的棲息環境。然而,僅僅依靠大型保護區是不夠的。台灣的保育專家們正在推動建立更多的小型保護區和生態廊道,特別是在都市和農業地區。例如,在台北市,一些社區正在實施「綠屋頂」計劃,這不僅為城市蜘蛛提供了新的棲息地,還有助於提高公眾對都市生物多樣性的認識。
其次,台灣正在積極推動生態友好型農業practices。這包括減少農藥使用、推廣有機耕作和實施綜合病蟲害管理。在台灣中部的某些茶園,農民們正在嘗試使用蜘蛛作為生物防治的一部分。通過減少農藥使用和保留田間的自然植被,這些茶園不僅維持了較高的蜘蛛多樣性,還實現了更加可持續的生產模式。這種方法不僅保護了蜘蛛,還利用了蜘蛛的生態服務功能,實現了雙贏。
針對外來入侵種的威脅,台灣正在加強入境檢疫和監測系統。同時,對於已經入侵的外來蜘蛛物種,如褐牆蜘蛛(Nesticodes rufipes),正在進行深入的生態學研究,以評估其對本土生態系統的影響,並制定適當的管理策略。在某些情況下,可能需要採取積極的移除措施,但這必須建立在充分的科學研究基礎之上,以避免造成新的生態問題。
台灣的蜘蛛保育策略還特別關注特有種和稀有種的保護。例如,台灣特有的黃條盲蛛(Haplogaleoids formosensis)正面臨棲息地喪失的威脅。為了保護這種蜘蛛,研究人員正在進行詳細的分布調查和生態需求研究,以便制定針對性的保護措施。這可能包括劃定特殊保護區,或者在其棲息地周邊實施更嚴格的土地使用管制。
公眾教育和參與是台灣蜘蛛保育策略中的重要組成部分。台灣的自然教育中心和博物館正在開展各種教育計劃,旨在提高公眾對蜘蛛的認識和欣賞。例如,台北市立動物園定期舉辦「蜘蛛觀察日」活動,邀請公眾參與蜘蛛多樣性調查。這不僅有助於收集寶貴的科學數據,還能夠培養公眾的生態意識。
在應對氣候變遷對蜘蛛的影響方面,台灣正在建立長期監測網絡。在全島不同海拔和生態區域設立固定監測點,定期進行蜘蛛群落調查。這些數據將有助於了解氣候變化如何影響蜘蛛的分布和行為,從而制定更有針對性的適應策略。例如,對於一些高山特有種蜘蛛,可能需要考慮建立人工輔助遷移計劃,以幫助它們適應不斷上升的溫度。
為了減少光污染對夜行性蜘蛛的影響,台灣的一些地方正在實施「暗空保護區」計劃。這些區域通過控制人工照明的強度和方向,為夜行性生物提供更好的活動環境。例如,在墾丁國家公園的某些區域,已經開始使用特殊設計的路燈,這些路燈能夠減少對周圍環境的光污染,同時仍然滿足人類活動的需求。
台灣的蜘蛛保育策略還包括加強法律保護。雖然目前台灣的野生動物保護法並未特別列出受保護的蜘蛛物種,但正在考慮將一些稀有和特有的蜘蛛物種納入保護名錄。這將為這些物種提供更強有力的法律保護,同時也有助於提高公眾對蜘蛛保護重要性的認識。
科學研究在台灣的蜘蛛保育策略中扮演著關鍵角色。台灣的研究機構正在加強對本土蜘蛛的基礎生態學研究,包括分類學、行為生態學和分子生物學等方面。這些研究不僅有助於發現和描述新物種,還能為保育決策提供科學依據。例如,通過分子生物學研究,科學家們正在揭示台灣特有蜘蛛的演化歷史和遺傳多樣性,這對於制定有效的保護策略至關重要。
此外,台灣正在推動跨領域合作,將蜘蛛保育與其他生態保護和可持續發展目標結合起來。例如,在都市規劃中,正在探索如何將蜘蛛棲息地需求納入綠地設計。在農業領域,正在研究如何將蜘蛛保護與有機農業和生態旅游相結合,創造經濟和生態的雙重效益。
台灣的蜘蛛保育策略還注重國際合作。通過參與國際會議和研究項目,台灣的科學家和保育工作者不僅分享了本土的經驗,還學習了其他地區的先進做法。這種國際交流有助於提升台灣蜘蛛保育工作的整體水平,同時也為全球生物多樣性保護做出貢獻。
綜上所述,台灣的蜘蛛保育策略是一個全面、多層次的體系,涵蓋了從基礎研究到實際保護措施的各個方面。這些策略不僅針對蜘蛛本身,還考慮了整個生態系統的健康和人類社會的需求,反映了現代保育理念的整體性和可持續性。通過這些努力,台灣希望能夠在保護其獨特的蜘蛛fauna的同時,也為全球生物多樣性保護做出貢獻。
13.3 棲息地喪失對蜘蛛的影響
棲息地喪失對台灣蜘蛛的影響是一個複雜而深遠的問題,涉及生態系統的多個層面。隨著台灣經濟的快速發展和人口增長,大規模的都市化和工業化進程不可避免地導致了自然棲息地的破壞和喪失。這種變化對蜘蛛群落造成了多方面的影響。
首先,棲息地喪失直接減少了蜘蛛的生存空間。許多蜘蛛物種對特定的環境條件有嚴格要求,例如某些特化的森林物種需要特定的樹種、濕度和光照條件。當這些環境被破壞時,這些蜘蛛物種就失去了賴以生存的家園。以台灣中部的八仙山自然保護區為例,近年來周邊地區的開發導致保護區面積縮小,一些只生活在原始林中的蜘蛛物種,如某些皿蛛科的物種,其分布範圍明顯減少。
其次,棲息地喪失導致蜘蛛群落結構發生改變。通常,棲息地喪失不僅僅是面積的減少,還伴隨著環境質量的下降和碎片化。這種變化往往有利於一些適應性強的廣布種,而不利於特化程度高的稀有種。例如,在台北市郊的一項研究中發現,隨著都市化程度的增加,原本多樣的蜘蛛群落逐漸被少數幾種適應城市環境的物種所主導,如一些常見的園蛛科物種。這種群落結構的簡化不僅降低了生物多樣性,還可能影響整個生態系統的穩定性。
棲息地喪失還會導致蜘蛛種群之間的隔離。當大片連續的棲息地被分割成小塊時,不同種群之間的基因交流就會受到阻礙。這種情況在台灣的山區尤為明顯,where道路和農田的開發將原本連續的森林切割成孤立的片段。例如,台灣特有的黃條盲蛛(Haplogaleoids formosensis)就面臨這樣的威脅。這種蜘蛛主要分布在中低海拔的森林中,棲息地的碎片化可能導致種群間的隔離,增加近親繁殖的風險,最終影響物種的長期生存。
棲息地喪失還會影響蜘蛛的食物來源。許多蜘蛛是重要的捕食者,它們的生存依賴於昆蟲和其他小型無脊椎動物。當棲息地被破壞時,這些獵物的數量和多樣性也會減少,從而影響蜘蛛的生存和繁殖。在台灣南部的某些農業地區,大規模的單一作物種植和過度使用農藥導致昆蟲多樣性下降,進而影響了當地蜘蛛的種群數量和組成。
此外,棲息地喪失還可能改變蜘蛛的行為模式。例如,一些原本在森林中生活的蜘蛛物種可能被迫適應都市環境。在台中市的一項研究中發現,某些原本棲息在樹林中的蜘蛛,如白額高腳蛛(Heteropoda venatoria),開始出現在建築物內部。這種行為改變雖然顯示了蜘蛛的適應能力,但也可能帶來新的生存挑戰,如與人類的衝突增加。
棲息地喪失對蜘蛛的影響還表現在其生態系統服務功能的減弱。蜘蛛作為重要的生物防治者,在控制害蟲方面發揮著關鍵作用。當棲息地喪失導致蜘蛛數量減少時,這種生態服務也會受到影響。在台灣的一些農業區域,研究人員發現,隨著自然植被的減少,農田中的蜘蛛多樣性和數量也相應下降,這可能導致農民不得不增加農藥使用,形成惡性循環。
值得注意的是,棲息地喪失對不同蜘蛛物種的影響並不均衡。一般來說,特化程度高、分布範圍狹窄的物種受到的影響更大。例如,台灣的一些洞穴蜘蛛,如某些微距蛛科的物種,由於其特殊的棲息環境要求,對棲息地的變化特別敏感。一旦其所依賴的洞穴環境被破壞,這些物種就面臨滅絕的風險。
棲息地喪失還可能加劇氣候變化對蜘蛛的影響。完整的棲息地能夠為蜘蛛提供緩衝,幫助它們應對溫度和濕度的變化。當棲息地被破壞或碎片化時,蜘蛛就失去了這種保護。在台灣的高山地區,研究人員發現,隨著氣溫上升,一些高山蜘蛛物種的分布範圍正在向更高海拔移動。然而,如果棲息地連續性被破壞,這種遷移就可能受到阻礙。
此外,棲息地喪失還可能影響蜘蛛的進化過程。長期的棲息地隔離可能導致不同種群之間產生遺傳分化,甚至形成新的物種。然而,如果棲息地喪失過快或過於嚴重,這種自然的演化過程可能被打斷,導致遺傳多樣性的喪失。
應對棲息地喪失對蜘蛛的影響,需要綜合考慮生態學、保護生物學和城市規劃等多個領域的知識。在台灣,一些積極的措施正在實施,如建立生態廊道、恢復退化棲息地、推廣生態友好型農業等。例如,在台南市的某些地區,正在嘗試將都市綠地系統與周邊的自然區域連接起來,為蜘蛛等小型動物提供移動和擴散的通道。
棲息地喪失對蜘蛛的影響是一個持續的過程,需要長期的監測和研究。台灣的科研機構正在進行系統的調查,以更好地理解這一問題的動態變化。這些研究不僅有助於評估保護措施的效果,還能為未來的保育策略提供科學依據。通過這些努力,希望能夠在滿足人類社會發展需求的同時,最大限度地減少對蜘蛛及其棲息地的負面影響。
14. 蜘蛛研究與未來展望
Ÿ 蜘蛛研究的現代技術
Ÿ 公民科學在蜘蛛研究中的應用
Ÿ 蜘蛛在生物科技中的應用
Ÿ 台灣蜘蛛研究的現況與挑戰
Ÿ 蜘蛛學的未來發展方向
蜘蛛研究領域正經歷著一場革命性的變革,這得益於現代科技的飛速發展和跨學科研究方法的廣泛應用。在台灣,蜘蛛研究不僅關注本土物種的保護和生態功能,還致力於將研究成果應用於生物科技和環境監測等領域。這一綜合性的研究方向不僅推動了蜘蛛學科的發展,還為解決諸如生物多樣性保護、生態系統服務評估等重大問題提供了新的視角和方法。
現代蜘蛛研究採用了多種先進技術,極大地提高了研究的精度和效率。分子生物學技術的應用使得研究人員能夠深入了解蜘蛛的遺傳多樣性和演化歷史。例如,在台灣特有的黃條盲蛛(Haplogaleoids formosensis)研究中,科學家們運用DNA條形碼技術和基因組測序,揭示了這個物種的遺傳結構和種群動態。這些信息不僅有助於理解台灣蜘蛛的演化過程,還為制定有效的保護策略提供了科學依據。
高速攝影和三維運動分析技術的應用,使研究人員能夠詳細記錄和分析蜘蛛的行為模式。在台灣國立中興大學的一項研究中,科學家們使用高速攝影技術研究了金園蛛(Nephila pilipes)的捕食行為。通過分析蜘蛛如何快速包裹獵物,研究人員不僅加深了對蜘蛛行為的理解,還為仿生機器人的設計提供了靈感。
微型感測器和遠程追蹤技術的應用,使得長期、連續監測蜘蛛的活動成為可能。在台灣的某些自然保護區,研究人員將微型GPS追蹤器安裝在一些大型蜘蛛物種身上,如 白額高腳蛛(Heteropoda venatoria),以了解它們的活動範圍和遷移模式。這些數據不僅有助於了解蜘蛛的生態需求,還為評估棲息地連通性和設計保護區網絡提供了重要依據。
環境DNA(eDNA)技術的應用為蜘蛛多樣性調查開闢了新途徑。研究人員可以通過分析水體或土壤樣本中的DNA片段來檢測蜘蛛的存在,這種方法特別適用於那些難以直接觀察的稀有或隱蔽性物種。在台灣的一些濕地生態系統中,科學家們正在使用eDNA技術來評估水生和半水生蜘蛛的分布情況,這為全面了解台灣的蜘蛛多樣性提供了新的工具。
人工智能和機器學習技術在蜘蛛研究中的應用也日益廣泛。例如,台灣的研究團隊開發了一種基於深度學習的蜘蛛識別系統,能夠通過分析蜘蛛網的結構特徵來識別不同的物種。這種技術不僅大大提高了蜘蛛調查的效率,還為公民科學項目提供了有力的支持。
公民科學在蜘蛛研究中的應用是另一個值得關注的趨勢。台灣的研究機構和保育組織正積極推動公眾參與蜘蛛研究和保護工作。例如,台灣特有生物研究保育中心發起的「蜘蛛觀察家」項目,邀請公眾通過智能手機APP上傳他們觀察到的蜘蛛照片和位置信息。這些數據經過專家審核後,被納入台灣蜘蛛分布數據庫,為研究人員提供了寶貴的長期、大尺度數據。
公民科學不僅有助於數據收集,還能提高公眾的環境意識。台北市立動物園定期舉辦的「蜘蛛生態體驗營」就是一個很好的例子。參與者不僅學習識別常見的蜘蛛物種,還參與簡單的生態調查活動。這種親身參與的經歷有助於消除公眾對蜘蛛的誤解和恐懼,培養他們對生物多樣性保護的興趣。
在生物科技領域,蜘蛛,特別是蛛絲,正成為研究的熱點。台灣的研究人員正在探索將蛛絲蛋白應用於醫療器械和生物材料領域。例如,國立台灣大學的一個研究團隊正在研究如何利用基因工程技術大規模生產蛛絲蛋白,並將其應用於製造高強度、生物相容性好的手術縫線。這種源於蜘蛛的生物材料有望在未來的醫療實踐中發揮重要作用。
蜘蛛毒素也是生物科技研究的重要對象。台灣的一些製藥公司正在研究如何利用蜘蛛毒素開發新型藥物。例如,從台灣捕鳥蛛(Psechrus taiwanensis)毒液中分離出的某些肽類化合物,被發現具有潛在的鎮痛作用,目前正在進行深入研究,有望開發成為新型止痛藥。
此外,蜘蛛在生物防治領域的應用也備受關注。台灣的一些有機農場正在探索如何利用本土蜘蛛物種來控制農業害蟲。例如,在南投縣的某些茶園,農民們通過保留田間植被和減少農藥使用,鼓勵蜘蛛自然繁殖,以此來控制茶樹害蟲。這種生態友好的害蟲管理方法不僅減少了化學農藥的使用,還有助於維持農田生態系統的平衡。
儘管台灣的蜘蛛研究取得了顯著進展,但仍面臨諸多挑戰。首先是資金和人力資源的限制。蜘蛛研究往往需要長期、持續的投入,但相對於其他生物學領域,蜘蛛研究獲得的資金支持較為有限。這導致一些長期生態監測項目難以為繼,影響了研究的連續性和深度。
其次是研究對象的特殊性帶來的困難。許多蜘蛛物種體型小、習性隱蔽,加上台灣複雜的地形,使得全面的物種調查和生態研究變得困難。特別是一些稀有物種或特有種,由於樣本量少,難以進行深入的生態學和行為學研究。
另一個挑戰是跨學科合作的需求。現代蜘蛛研究越來越依賴於多學科的合作,如生態學、分子生物學、信息科學等。然而,有時候不同學科之間的溝通和合作存在障礙,這可能會影響研究的進展和成果的應用。
此外,氣候變化和人類活動對蜘蛛棲息地的影響,也給研究工作帶來了新的挑戰。研究人員需要不斷調整研究方法和重點,以應對這些快速變化的環境條件。
面對這些挑戰,台灣的蜘蛛研究正在積極調整策略和方向。首先是加強基礎研究和應用研究的結合。例如,在研究台灣特有蜘蛛物種的同時,也探索這些物種在生物防治或生物材料方面的潛在應用價值。這種結合不僅有助於吸引更多的研究資金,還能提高研究成果的社會影響力。
其次是推動跨學科合作和國際交流。台灣的研究機構正在建立跨學科研究平台,鼓勵生態學家、分子生物學家、材料科學家等共同參與蜘蛛研究。同時,也在積極參與國際合作項目,如東亞蜘蛛多樣性調查計劃,這不僅有助於提升台灣蜘蛛研究的國際影響力,還能借鑒其他國家和地區的先進經驗。
第三是加強公眾參與和科普教育。通過開展更多的公民科學項目和科普活動,不僅能夠提高公眾對蜘蛛的認識和保護意識,還能吸引更多年輕人加入蜘蛛研究領域。例如,台灣自然科學博物館正在籌劃一個大型的蜘蛛生態展,通過互動展品和虛擬實境技術,讓公眾近距離體驗蜘蛛的奇妙世界。
第四是發展新的研究方法和技術。例如,利用無人機技術進行大尺度的蜘蛛網分布調查,或者開發新的分子標記技術來研究蜘蛛的種群遺傳結構。這些新技術的應用有望突破傳統研究方法的局限,為蜘蛛研究開闢新的領域。
在未來,台灣的蜘蛛研究有望在以下幾個方向取得突破:
首先是在物種多樣性和系統分類學方面。隨著分子生物學技術的進步和野外調查的深入,預計將會發現更多的新物種,特別是一些隱蔽性強的小型蜘蛛。同時,對已知物種的分類地位也可能會有新的認識,這將有助於更好地理解台灣蜘蛛fauna的演化歷史和生物地理學特徵。
其次是在生態功能和生態系統服務方面的研究。隨著生態學理論和研究方法的發展,研究人員將能夠更精確地量化蜘蛛在生態系統中的作用,例如它們在控制害蟲、促進養分循環等方面的具體貢獻。這些研究將為生態系統管理和生態恢復提供重要的科學依據。
第三是在行為生態學和認知科學領域。隨著神經生物學和行為分析技術的進步,研究人員有望深入了解蜘蛛的感知世界和決策機制。例如,探索蜘蛛如何感知和應對環境變化,或者它們如何在複雜的三維空間中進行導航。這些研究不僅有助於理解蜘蛛的生存策略,還可能為人工智能和機器人技術提供新的靈感。
第四是在生物材料和生物醫學領域的應用研究。隨著合成生物學和材料科學的發展,有望實現蛛絲蛋白的大規模生產和定制化設計。這將為開發新型的高性能纖維、醫用材料等提供可能。同時,蜘蛛毒素研究也可能為新藥開發開闢新的途徑,特別是在神經系統疾病和疼痛管理領域。
第五是在全球變化生物學領域。蜘蛛由於其廣泛分布和對環境變化的敏感性,成為研究氣候變化和人類活動影響的理想對象。長期、大尺度的蜘蛛群落監測數據將有助於預測和評估生態系統對全球變化的響應,為制定適應和緩解策略提供科學依據。
此外,蜘蛛研究還可能在生物仿生學、都市生態學、保育生物學等多個領域發揮重要作用。例如,研究蜘蛛如何在城市環境中適應和生存,可能為設計更加生態友好的都市環境提供參考。
台灣的蜘蛛研究正處於一個充滿機遇和挑戰的時期。隨著研究方法的創新和跨學科合作的深入,蜘蛛研究有望在生態保護、生物技術、醫藥開發等多個領域做出重要貢獻。同時,這些研究也將加深我們對這些神奇生物的理解,提高公眾的環境意識,最終推動人類與自然的和諧共處。
台灣作為一個生物多樣性熱點,其蜘蛛研究不僅對本地生態保護具有重要意義,還可能為全球蜘蛛學研究提供獨特的洞見和案例。通過持續的投入和創新,台灣的蜘蛛研究有望在國際蜘蛛學界佔據重要地位,為解決全球性的生態環境問題貢獻智慧和力量。
在台灣蜘蛛研究的未來發展中,跨領域整合將成為一個重要趨勢。例如,將蜘蛛生態學研究與景觀生態學、保育生物學相結合,可以更好地理解棲息地破碎化對蜘蛛群落的影響。在墾丁國家公園的一項研究中,科學家們正在利用地理信息系統(GIS)技術,結合蜘蛛分布數據和景觀特徵,分析棲息地連通性對蜘蛛多樣性的影響。這種跨學科的方法不僅有助於制定更有效的保護策略,還為理解生態系統功能提供了新的視角。
另一個值得關注的方向是蜘蛛在環境監測中的應用。由於蜘蛛對環境變化敏感,它們可以作為生態系統健康的指示物種。台灣的研究人員正在探索利用特定蜘蛛物種的存在或缺失來評估生態系統的質量。例如,在台中市的都市綠地研究中,科學家們發現某些蜘蛛物種的分布與空氣質量和綠地連通性有密切關係。這種研究不僅為城市規劃提供了新的工具,還有助於提高公眾對城市生物多樣性的認識。
在生物防治領域,台灣的蜘蛛研究也在不斷深入。除了傳統的害蟲控制應用,研究人員還在探索蜘蛛在控制外來入侵物種方面的潛力。例如,在應對近年來困擾台灣農業的秋行軍蟲(Spodoptera frugiperda)問題時,科學家們正在研究本土蜘蛛是否能成為有效的天敵。這種研究不僅有助於減少農藥使用,還可能為生態平衡管理提供新的思路。
蜘蛛毒素研究是另一個快速發展的領域。台灣的研究團隊正在利用先進的蛋白質組學和藥理學技術,深入研究蜘蛛毒素的結構和功能。例如,從台灣鳥蛛(Macrothele gigas)毒液中分離出的某些神經毒素,被發現具有高度的離子通道特異性,這為開發新型疼痛治療藥物和神經系統疾病治療方案提供了可能。這種研究不僅有望帶來醫學突破,還可能為台灣的生物科技產業注入新的活力。
在蛛絲研究方面,台灣的科學家正在探索將蛛絲蛋白與其他材料結合,開發新型複合材料。例如,國立清華大學的一個研究團隊正在研究如何將蛛絲蛋白與碳納米管結合,以製造超高強度、超輕質的材料。這種材料可能在航空航天、運動器材等領域找到應用。同時,研究人員也在探索利用基因工程技術改造絲蛋白的性質,以適應不同的應用需求。
公民科學在台灣蜘蛛研究中的作用將進一步擴大。除了現有的觀察記錄項目,研究機構正在開發更加互動和遊戲化的參與方式。例如,一款名為「蜘蛛獵人」的手機遊戲正在開發中,玩家通過拍攝和上傳蜘蛛照片來獲得積分,同時這些數據也被納入科學研究數據庫。這種創新的方法不僅能吸引更多人參與科學研究,還能提高公眾的生態意識。
在蜘蛛行為學研究方面,台灣的科學家正在利用先進的三維動作捕捉技術和計算機視覺算法,深入分析蜘蛛的運動模式和社會行為。例如,在研究台灣金絲蛛(Nephila pilipes)的求偶行為時,研究人員使用高速攝像機和人工智能算法,精確追踪和分析蜘蛛的微小動作。這種研究不僅加深了我們對蜘蛛行為的理解,還可能為開發新型機器人提供靈感。
隨著環境DNA技術的成熟,台灣的研究人員正在探索將這種技術應用於大尺度蜘蛛多樣性調查。例如,在台灣東部的某些河流系統中,科學家們正在通過分析水樣中的DNA來檢測稀有或難以直接觀察的蜘蛛物種。這種方法不僅能提高調查效率,還可能發現一些以前未知的物種。
在蜘蛛與微生物相互作用的研究方面,台灣的科學家正在揭示一些令人驚訝的發現。例如,研究人員發現某些蜘蛛物種體表存在特殊的共生細菌,這些細菌可能幫助蜘蛛抵抗病原體。這項研究不僅開闢了理解蜘蛛免疫機制的新途徑,還可能為開發新型抗生素提供線索。
台灣的蜘蛛研究還在探索蜘蛛在文化和藝術領域的價值。例如,一些研究人員正在與藝術家合作,探索如何將蜘蛛網的結構美學應用於建築設計和視覺藝術創作。這種跨界合作不僅能激發創新思維,還能通過藝術形式提高公眾對蜘蛛的興趣和理解。
在教育領域,虛擬現實(VR)和增強現實(AR)技術正被用於開發互動式的蜘蛛教育工具。例如,台灣科學教育館正在開發一個VR蜘蛛生態系統,讓學生能夠虛擬體驗蜘蛛的日常生活和生態角色。這種創新的教育方式不僅能提高學習效果,還能培養學生的生態意識。
隨著基因編輯技術的發展,台灣的研究人員也在探索利用CRISPR技術研究蜘蛛基因功能。例如,通過基因編輯技術,科學家們希望能夠深入了解控制蛛絲產生的基因網絡,這可能為人工合成高性能蛛絲材料鋪平道路。
總的來說,台灣的蜘蛛研究正朝著更加多元化、跨學科和應用導向的方向發展。這些研究不僅有助於加深我們對這些神奇生物的理解,還可能為解決諸如生物多樣性保護、新材料開發、藥物研發等重大問題提供新的思路和方法。通過持續的創新和努力,台灣的蜘蛛研究有望在國際舞台上發揮更大的影響力,為全球生態保護和科技創新做出獨特貢獻。