永續社 www.epa.url.tw 林雨莊
生命的歷史可以追溯到約38億年前,當時地球的環境尚處於原始狀態,充滿了有毒氣體和高溫高壓的條件。在這樣的環境下,生命的最早形式開始出現。這些原始的生物體被稱為原核生物,與今日的細菌和古菌相似。它們是單細胞的,沒有明確的細胞核或其他膜狀胞器。這些原核生物通過簡單的代謝方式,尤其是厭氧代謝,來獲取能量。這個時期稱為地質史上的冥古宙(Hadean eon)。
隨著時間的推移,地球環境逐漸穩定,早期生物的演化也逐漸進入一個關鍵階段。大約35億年前,出現了光合作用的原核生物,這些生物能夠利用陽光將水和二氧化碳轉化為氧氣和葡萄糖,這一過程對地球的生態系統產生了深遠的影響。這種氧氣的累積最終導致大氣中的氧含量逐漸增多,並形成了地球的臭氧層,為後來更複雜的生命形式提供了適合的環境。
隨著氧氣的增加,生命開始進入一個新的演化階段。大約20億年前,真核生物出現,這些生物具有細胞核和膜狀胞器。真核生物的出現代表著生命結構上的一大飛躍,因為它們能夠進行更複雜的代謝活動,並具有更強的適應性。這一時期被稱為原生代(Proterozoic eon)。真核生物的演化還導致了多細胞生物的出現,這些生物的細胞能夠分化並執行不同的功能,這為後來動植物的出現奠定了基礎。
到了約5.4億年前的寒武紀(Cambrian period),生命形式的多樣性達到了一個顯著的高峰,這一時期的生物爆發性增長,被稱為寒武紀大爆發。在此期間,出現了多種多樣的動物門類,這些動物擁有不同的身體結構和適應策略。許多現代動物門的祖先都可以追溯到這個時期,這包括軟體動物、節肢動物和脊椎動物。這一時期的生物以海洋為主要生活環境,並開始形成完整的食物鏈和生態系統。
隨著時間推進,生命從海洋逐漸登陸。這一過程發生在約4億年前的泥盆紀(Devonian period),當時一些原始的植物和無脊椎動物開始適應陸地生活。這些早期的陸生植物主要是蕨類植物,它們通過發展根系和維管束系統來吸收水分和養分。動物方面,最早的四足類動物(四肢動物的祖先)也在這一時期出現,它們的出現象徵著動物正式從水生環境向陸地環境的過渡。
接下來的石炭紀(Carboniferous period)是地球植被和生物多樣性急劇增長的時期,這一時期的巨大森林為後來的煤層形成提供了物質基礎。同時,兩棲類和爬行類動物在這段時間進一步繁盛,並逐漸向陸地生態系統的各個角落擴散。
隨後的二疊紀(Permian period)則是古生代的尾聲,這一時期標誌著地球上一次大型生物滅絕事件的發生。這場滅絕事件導致了海洋和陸地上多達90%以上的物種滅亡,但也為恐龍等新型生物的崛起鋪平了道路。
中生代(Mesozoic era)是恐龍的時代,它們在三疊紀(Triassic period)至白堊紀(Cretaceous period)之間主宰了地球的生態系統。同時,第一批鳥類和哺乳動物也在這一時期出現。白堊紀末期,一場隕石撞擊地球的事件導致了另一場生物大滅絕,恐龍等大型爬行動物滅絕,而哺乳動物則在這場災難後迅速崛起。
進入新生代(Cenozoic era),哺乳動物和鳥類成為了地球生態系統中的主要角色,這一時期的氣候變遷和地理變化促進了它們的進一步多樣化。人類的祖先出現在約600萬年前的非洲,並在後來的數百萬年間逐漸演化為智人(Homo sapiens),最終成為了地球上最具影響力的生物。
生命的歷史是一個不斷演化的過程,充滿了創造和毀滅、適應和變遷。從原始的單細胞生物到今日複雜的多細胞生物,生命的多樣性和適應能力無疑是地球上最為奇妙的現象之一。這個漫長的演化過程展示了生命在面對環境變遷和生態壓力時的強大生存能力,也揭示了地球本身作為生命孕育之地的獨特性。
生命的歷史是生物學中一個極其重要且引人入勝的領域。它不僅揭示了地球上生命的起源和演化過程,還幫助我們理解現今生物的多樣性和複雜性。研究生命的歷史對於我們理解自身的起源、與其他生物的關係,以及我們在地球生態系統中的角色都具有重要意義。
生命歷史的研究涵蓋了從最早的單細胞生物到現代複雜生物的漫長演化過程。這個過程橫跨數十億年,涉及無數的物種和環境變遷。通過研究化石記錄、比較現存物種的DNA,以及分析地質證據,科學家們逐漸拼湊出了生命演化的宏大圖景。這些研究不僅幫助我們了解過去,還為預測未來生物演化提供了寶貴的洞見。
了解生命的歷史對於醫學和公共衛生領域也有重要意義。通過研究病原體的演化史,科學家們可以更好地理解疾病的傳播機制和抗藥性的發展。這些知識對於開發新的治療方法和預防策略至關重要。例如,了解病毒的演化歷史有助於預測未來可能出現的新型病毒,從而更好地應對全球性的健康威脅。
生命歷史的研究還為保護生物多樣性提供了重要依據。通過了解物種如何適應不同的環境,以及過去的滅絕事件是如何發生的,我們可以制定更有效的保護策略。這對於應對當前的生態危機,如氣候變化和棲息地破壕,具有重要意義。保護生物多樣性不僅對維持生態平衡至關重要,還可能為人類提供新的藥物來源和其他重要資源。
從哲學和文化的角度來看,生命的歷史也具有深遠的影響。它挑戰了人類中心主義的觀點,讓我們認識到人類只是地球生命長河中的一個分支。這種認識有助於培養對其他生物的尊重和對生態系統的珍惜。同時,了解生命的歷史也激發了人們對宇宙中其他可能存在生命的探索興趣,推動了天文生物學等新興領域的發展。
在教育領域,生命的歷史是激發學生對科學興趣的重要途徑。通過學習生命演化的過程,學生們可以培養批判性思維和科學推理能力。同時,這也有助於培養學生的環境意識和可持續發展理念。
生命歷史的研究還為多個科技領域提供了靈感和指導。例如,仿生學(Biomimetics)借鑒生物在長期演化中形成的結構和功能,開發新的材料和技術。這種跨學科的應用展現了生命歷史研究的廣泛影響力。
此外,了解生命的歷史對於應對全球性挑戰也具有重要意義。氣候變化是當前人類面臨的最大威脅之一,而通過研究過去的氣候變化如何影響生物演化,我們可以更好地預測和應對未來的氣候危機。同樣,在糧食安全方面,了解作物的演化歷史有助於開發更具抗性和產量的新品種。
生命歷史的研究還揭示了生命的韌性和適應能力。地球歷史上曾經發生過多次大規模滅絕事件,但生命總是能夠以新的形式重新繁衍。這種洞察不僅給予我們面對當前環境挑戰的希望,也為探索極端環境中的生命可能性提供了理論基礎。
在社會科學領域,生命的歷史研究也有重要應用。例如,在法律和倫理領域,對生命演化的理解影響了我們對物種保護、動物權利等問題的看法。在經濟學中,生態系統服務的價值評估也越來越受到重視,這在很大程度上基於對生態系統演化歷史的理解。
總的來說,生命的歷史是一個涉及多學科、影響深遠的研究領域。它不僅幫助我們理解過去,還為塑造更美好的未來提供了重要指導。無論是在科學研究、技術創新,還是在社會發展和環境保護方面,對生命歷史的深入理解都將繼續發揮關鍵作用。
進化是生物學中最核心和最具革命性的概念之一,它解釋了地球上生命的多樣性和複雜性。進化理論的核心思想是,生物隨著時間的推移而改變,這種改變是通過遺傳變異和自然選擇的過程實現的。這個概念最初由查爾斯·達爾文在19世紀提出,並在之後的一個多世紀裡得到了大量證據的支持和完善。
進化的基本機制是遺傳變異和自然選擇的相互作用。遺傳變異是指在一個物種的個體之間存在的遺傳差異。這些差異可能源於基因突變、基因重組或其他遺傳機制。自然選擇則是指那些具有有利特徵的個體更有可能在其環境中生存和繁衍後代。隨著時間的推移,這些有利特徵在種群中變得更加普遍,從而導致物種的逐漸改變。
達爾文提出的自然選擇理論是建立在幾個關鍵觀察之上的。首先,他注意到所有物種都有產生比環境所能維持的更多後代的潛力。其次,他觀察到同一物種的個體之間存在變異。第三,他認識到這些變異中的一些是可以遺傳的。最後,他推斷那些最適應其環境的個體更有可能生存和繁衍。這些觀察和推理形成了自然選擇理論的基礎。
進化不僅僅是一個歷史過程,它在當今世界中仍在持續發生。我們可以在許多方面觀察到進化的證據,如細菌對抗生素的抗藥性、農作物害蟲對殺蟲劑的抗性,以及病毒的快速變異。這些例子展示了進化如何在相對短的時間內發生,並對人類社會產生直接影響。
化石記錄是支持進化理論的最強有力的證據之一。化石為我們提供了生物隨時間變化的直接證據,讓我們能夠追溯物種的演化歷程。例如,馬的化石記錄清楚地展示了從小型多趾動物到現代大型單蹄馬的漸進式變化。此外,過渡性化石,如始祖鳥(Archaeopteryx),為我們理解主要生物類群之間的演化關係提供了重要線索。
比較解剖學也為進化理論提供了有力支持。不同物種之間的結構相似性,特別是同源結構的存在,表明它們可能有共同的祖先。例如,所有哺乳動物的前肢骨骼結構基本相似,無論是用於飛行、游泳還是奔跑,這表明它們源自共同的祖先形式。同樣,痕跡器官的存在,如人類的闌尾或鯨魚的髖骨殘跡,也為進化提供了證據。
分子生物學的發展為進化研究提供了新的強大工具。DNA序列分析使科學家能夠精確地測量不同物種之間的遺傳相似性,從而重建演化關係。這種方法不僅確認了基於形態學的許多演化關係,還揭示了一些意想不到的聯繫。例如,分子研究顯示鯨類與河馬有近似的演化關係,這是純粹基於形態學難以發現的。
自然選擇並不是進化的唯一機制。遺傳漂變(Genetic drift)是另一個重要的進化力量,特別是在小型種群中。遺傳漂變是指基因頻率的隨機變化,這種變化不依賴於特定等位基因(Allele)的適應性優勢。在小種群中,這種隨機變化可能導致某些等位基因的固定或丟失,從而改變種群的遺傳構成。
基因流動(Gene flow)是指不同種群之間的基因交換,通常通過個體的遷移或花粉的傳播實現。基因流動可以增加種群的遺傳變異,但也可能阻礙局部適應。例如,如果兩個適應不同環境的種群之間存在持續的基因流動,可能會阻礙它們向不同物種分化的過程。
突變是遺傳變異的最終來源,也是進化的關鍵驅動力之一。突變可以是有害的、中性的或有利的。雖然大多數突變可能是有害的或中性的,但偶爾出現的有利突變可能會被自然選擇保留並在種群中傳播。隨著時間的推移,這些突變的累積可以導致顯著的進化變化。
適應是進化過程的一個核心概念,指的是生物特徵如何使有機體更好地適應其環境。適應可以涉及形態、生理或行為的變化。例如,仙人掌的特殊結構是對乾旱環境的適應,而北極熊的白色皮毛則是對其雪地環境的適應。然而,重要的是要認識到不是所有的特徵都是適應性的,有些可能是歷史遺留或其他進化過程的副產品。
協同進化(Coevolution)是指兩個或多個物種之間的相互適應過程。這種現象在捕食者-獵物關係、寄生關係和共生關係中特別常見。例如,花朵和授粉昆蟲之間的協同進化導致了高度專門化的互利關係,如某些蘭花與特定蛾類之間的關係。
物種形成是進化過程的一個關鍵方面,它解釋了新物種如何產生。物種形成通常發生在種群被地理障礙分隔(異域物種形成)或生態位分化(同域物種形成)的情況下。隨著時間的推移,這些分離的種群可能會積累足夠的遺傳差異,以至於不能再相互交配,從而形成新的物種。
進化理論不僅解釋了生物多樣性的起源,還為我們理解許多生物現象提供了框架。例如,它解釋了為什麼生物體不是完美的,而是充滿了妥協和權衡。這是因為進化是在現有結構的基礎上工作,而不是從頭開始設計。
進化的概念也擴展到了生物學之外的領域。在醫學中,進化思想幫助我們理解疾病的起源和傳播,以及抗藥性的產生。在農業中,它指導了作物和牲畜的育種策略。在計算機科學中,進化算法被用來解決複雜的優化問題。
值得注意的是,雖然進化是一個科學事實,得到了大量證據的支持,但它在某些社會中仍然存在爭議。這主要是由於對進化理論的誤解,或者它與某些宗教信仰的表面衝突。然而,許多宗教領袖和學者認為,進化理論與宗教信仰並不必然矛盾。
理解進化不僅對科學研究至關重要,對於公眾科學素養也同樣重要。它幫助我們認識到生命的相互聯繫,理解人類在自然界中的位置,並為我們面對諸如氣候變化、生物多樣性喪失等全球挑戰提供了重要的視角。
自然選擇是進化理論的核心機制,由查爾斯·達爾文在19世紀提出。它解釋了生物如何適應其環境,以及物種如何隨時間變化。自然選擇的基本原理是,那些最適應環境的個體更有可能生存和繁衍後代,從而將其有利特徵傳給下一代。這個過程導致了物種的漸進式改變,最終可能導致新物種的形成。
自然選擇的運作基於幾個關鍵概念。首先是變異(Variation)的存在。在任何給定的種群中,個體之間都存在差異。這些差異可能涉及形態、生理或行為特徵。例如,在一群斑馬中,每個個體的條紋模式略有不同。這些差異部分是由遺傳因素決定的,這意味著它們可以傳給後代。
第二個關鍵概念是過度繁殖(Overproduction)。大多數物種產生的後代數量遠超過環境所能支持的數量。例如,一棵橡樹在其一生中可能產生數十萬個橡子,但只有極少數會成長為成熟的樹木。這種過度繁殖導致了生存競爭。
第三個概念是適應度(Fitness)。在生物學中,適應度指的是一個個體在特定環境中生存和繁殖的能力。那些具有有利特徵的個體往往有更高的適應度。例如,在寒冷環境中,具有更厚皮毛的動物可能比同種的薄毛個體有更高的生存機會。
自然選擇的過程可以概括為以下幾個步驟:首先,種群中存在遺傳變異。然後,由於資源有限和過度繁殖,會產生生存競爭。那些具有有利特徵的個體更有可能在這種競爭中生存下來。這些倖存的個體更有可能繁衍後代,並將其有利特徵傳給下一代。隨著時間的推移,這些有利特徵在種群中變得更加普遍。
自然選擇可以導致不同類型的適應。趨向選擇(Directional selection)favors one extreme of a trait。例如,在長頸鹿的進化過程中,更長的脖子可能被選擇,因為它允許動物接觸到更高的食物來源。穩定選擇(Stabilizing selection)favors the average individuals in a population。例如,人類嬰兒的出生體重受到穩定選擇的影響,因為太輕或太重的嬰兒都有較高的死亡風險。分裂選擇(Disruptive selection)favors both extremes of a trait。這可能導致種群分化為兩個不同的亞種。
自然選擇不僅作用於明顯的外部特徵,也影響著生理和行為特徵。例如,抗生素耐藥性細菌的出現就是自然選擇的結果。在抗生素的存在下,那些偶然擁有抗藥性基因的細菌更有可能存活和繁殖,導致抗藥性在細菌種群中迅速擴散。
性選擇(Sexual selection)是自然選擇的一種特殊形式,它涉及到個體在獲得配偶方面的競爭。這可能導致一些看似與生存無關,甚至可能有害的特徵的進化。孔雀的華麗尾羽就是一個典型的例子。雖然這些尾羽可能使孔雀更容易被捕食者發現,但它們在吸引雌性方面非常有效,因此被性選擇保留了下來。
自然選擇並不總是導致完美的適應。這是因為自然選擇只能在現有變異的基礎上工作,而不能憑空創造新的特徵。此外,環境不斷變化,使得完美的適應變得困難。例如,人類的脊椎雖然適應了直立行走,但並不完美,這就是為什麼許多人會遭受背痛。
重要的是要理解,自然選擇作用於個體,但其效果在種群水準上表現出來。個體不會在其一生中進化,而是種群隨著世代更替而改變。此外,自然選擇不具有目的性或方向性。它只是保留了在當前環境中最有利的特徵,而不考慮這些特徵在未來是否仍然有利。
自然選擇的速度可能因情況而異。在某些情況下,如細菌對抗生素的適應,變化可能非常迅速。在其他情況下,如大型動物的形態變化,可能需要數百萬年。環境壓力的強度、種群大小、世代時間等因素都會影響自然選擇的速度。
自然選擇並不是進化的唯一機制。遺傳漂變、基因流動和突變等過程也在物種的變化中發揮作用。然而,自然選擇是唯一能夠產生適應性變化的機制,這使得它在進化理論中具有核心地位。
自然選擇的概念不僅在生物學中重要,也已經擴展到其他領域。在醫學中,理解自然選擇有助於我們應對抗生素耐藥性和病毒的快速進化。在農業中,它指導了作物和牲畜的育種策略。在計算機科學中,模擬自然選擇的遺傳算法被用於解決複雜的優化問題。
然而,自然選擇的概念有時會被誤解或濫用。例如,社會達爾文主義試圖將自然選擇的概念不當地應用於人類社會,這導致了一些有害的社會政策。重要的是要認識到,自然選擇是一個生物學過程,不應被用來證明任何社會或政治意識形態。
自然選擇也常常被誤解為一個有目的的過程,彷彿自然在"選擇"最好的個體。實際上,自然選擇只是那些恰好擁有有利特徵的個體更有可能生存和繁衍的結果。它不涉及任何預見或計劃。
理解自然選擇對於理解生物多樣性至關重要。地球上的每個物種都是數十億年自然選擇過程的產物。即使是最簡單的生物體也顯示出驚人的適應性,這反映了自然選擇的強大力量。
自然選擇也幫助我們理解為什麼某些特徵在不同的物種中反複出現。例如,魚類和海豚雖然在進化上相距甚遠,但都演化出了流線型的身體,這是對水生環境的共同適應。這種現象被稱為趨同進化(Convergent evolution)。
在保護生物學中,理解自然選擇也很重要。當我們試圖保護瀕危物種時,我們需要考慮這些物種如何適應其環境,以及環境變化如何影響它們的生存機會。此外,了解自然選擇可以幫助我們預測物種如何應對氣候變化等全球性威脅。
自然選擇的概念也影響了我們對人類自身的理解。它解釋了我們的許多生物特徵,如直立行走、大腦大小、社會行為等。然而,重要的是要記住,文化和技術在人類進化中也扮演了重要角色,這些因素可能會改變或減弱自然選擇的影響。
進化論是生物學中最基本和最重要的理論之一,它解釋了地球上生命的多樣性和複雜性是如何隨時間演變而來的。這個理論的核心概念是,生物體通過世代相傳的遺傳變異和自然選擇的過程而逐漸改變。進化論不僅解釋了現存物種的起源,還揭示了所有生命形式之間的關聯。
進化論的發展歷程長達數個世紀,但其現代形式主要歸功於19世紀的英國博物學家查爾斯·達爾文。1859年,達爾文在其著作《物種起源》中首次全面闡述了進化理論。達爾文的理論建立在幾個關鍵觀察之上:首先,他注意到同一物種的個體之間存在變異;其次,這些變異中的一些是可以遺傳的;再次,生物體產生的後代數量遠超過能夠存活的數量;最後,那些最適應環境的個體更有可能存活和繁衍。
達爾文提出的自然選擇機制是進化論的核心。自然選擇的基本原理是,那些具有有利特徵的個體更有可能在競爭中生存下來並繁衍後代,從而將這些有利特徵傳給下一代。隨著時間的推移,這些有利特徵在種群中變得越來越普遍,導致物種的漸進式變化。
然而,達爾文的理論在當時還缺乏對遺傳機制的理解。這個缺口在20世紀初由孟德爾遺傳學的重新發現而得到填補。孟德爾的工作為理解遺傳變異如何在世代間傳遞提供了機制。這種綜合導致了現代綜合理論(Modern Synthesis)的形成,這個理論整合了達爾文的自然選擇理論和孟德爾的遺傳學。
現代進化論認為,進化的主要機制包括自然選擇、遺傳漂變、突變和基因流動。自然選擇是產生適應性變化的唯一機制,而其他機制則可能導致中性或非適應性的變化。突變是新遺傳變異的最終來源,它為自然選擇提供了原材料。遺傳漂變是指基因頻率的隨機變化,特別是在小種群中可能產生顯著影響。基因流動則是指不同種群之間的基因交換。
進化論的一個重要方面是它解釋了生物多樣性的起源。根據這個理論,所有現存的物種都源自共同的祖先,通過漫長的進化過程分化而來。這個過程稱為物種形成,通常發生在種群被地理障礙分隔(異域物種形成)或生態位分化(同域物種形成)的情況下。隨著時間的推移,這些分離的種群可能會積累足夠的遺傳差異,以至於不能再相互交配,從而形成新的物種。
進化論還解釋了為什麼生物體顯示出一定程度的適應性。適應是指生物特徵如何使有機體更好地適應其環境。例如,沙漠植物的特殊結構是對乾旱環境的適應,而極地動物的厚皮毛則是對寒冷氣候的適應。然而,進化論也解釋了為什麼適應並不完美。這是因為進化只能在現有變異的基礎上工作,而且環境不斷變化,使得完美適應變得困難。
化石記錄為進化論提供了強有力的支持。化石顯示了生物形態如何隨時間變化,並提供了物種出現和滅絕的證據。特別是過渡性化石,如始祖鳥(Archaeopteryx),為我們理解主要生物類群之間的演化關係提供了重要線索。
比較解剖學和胚胎學也為進化論提供了證據。不同物種之間的結構相似性,特別是同源結構的存在,表明它們可能有共同的祖先。例如,所有脊椎動物的胚胎在早期發育階段都顯示出驚人的相似性,這反映了它們的共同進化歷史。
分子生物學的發展為進化論提供了新的支持和洞見。DNA序列分析使科學家能夠精確地測量不同物種之間的遺傳相似性,並重建演化關係。這種方法不僅確認了基於形態學的許多演化關係,還揭示了一些意想不到的聯繫。例如,分子研究顯示鯨類與偶蹄類動物(如牛和羊)有近親關係。
進化論的一個重要應用是在醫學領域。理解進化有助於我們應對諸如抗生素耐藥性和病毒快速變異等問題。例如,流感病毒的快速進化解釋了為什麼我們需要每年更新流感疫苗。同樣,了解癌細胞如何進化也有助於開發更有效的治療策略。
在農業中,進化理論指導了作物和牲畜的育種策略。通過理解自然選擇的原理,農業科學家能夠開發出產量更高、抗病能力更強的作物品種。同時,進化理論也幫助我們理解和應對農業害蟲的抗藥性問題。
進化論還為我們理解人類自身提供了框架。它解釋了我們的生物學特徵,如直立行走、大腦大小、社會行為等是如何演化而來的。同時,它也幫助我們理解人類遺傳疾病的起源和分布。然而,重要的是要認識到,文化和技術在人類近期進化中扮演了重要角色,這些因素可能會改變或減弱自然選擇的影響。
儘管進化論在科學界已經得到廣泛接受,但它在某些社會群體中仍然存在爭議。這主要是由於對進化理論的誤解,或者它與某些宗教信仰的表面衝突。然而,許多宗教領袖和學者認為,進化理論與宗教信仰並不必然矛盾。重要的是要理解,進化論是一個科學理論,它解釋自然現象,而不涉及超自然或哲學問題。
進化論的影響遠遠超出了生物學領域。它改變了我們對自然界的理解,影響了從心理學到經濟學等多個學科。在哲學上,它挑戰了人類中心主義的觀點,讓我們認識到人類只是生命演化長河中的一個分支。在環境保護方面,進化理論為我們理解生態系統的複雜性和脆弱性提供了基礎,強調了保護生物多樣性的重要性。
理解進化論對於公眾科學素養也至關重要。它不僅幫助人們更好地理解科學新聞和政策決策,還培養了對自然世界的欣賞和尊重。在面對全球性挑戰如氣候變化時,進化論提供了理解生物如何應對環境變化的框架,這對於預測和緩解這些變化的影響至關重要。
進化理論是現代生物學的基石,它解釋了地球上生命的多樣性和複雜性是如何隨時間演變而來的。這個理論得到了來自多個科學領域的大量證據支持。這些證據不僅證實了進化的發生,還揭示了進化的機制和過程。以下我們將詳細探討支持進化理論的主要證據來源。
化石記錄是支持進化理論最直接和最有力的證據之一。化石為我們提供了生物隨時間變化的直接證據,讓我們能夠追溯物種的演化歷程。通過研究不同地質年代的化石,科學家們能夠重建生物形態的漸進式變化。例如,馬的化石記錄清楚地展示了從小型多趾動物到現代大型單蹄馬的漸進式變化。這個序列包括了原始的多趾馬科動物,如始祖馬(Hyracotherium),到後來的中間形式,如中新馬(Merychippus),最後到現代的馬屬(Equus)。
特別值得注意的是過渡性化石,它們展示了主要生物類群之間的中間特徵。始祖鳥(Archaeopteryx)就是一個著名的例子,它具有爬行動物和鳥類的特徵,為爬行動物向鳥類演化提供了重要證據。類似地,蒂克塔利克魚(Tiktaalik)是一種介於魚類和四足動物之間的過渡形式,它具有魚類的鰭,但同時也有原始的腿骨結構,為魚類向陸地脊椎動物的演化提供了重要線索。
比較解剖學為進化理論提供了另一個重要的證據來源。不同物種之間的結構相似性,特別是同源結構的存在,表明它們可能有共同的祖先。同源結構是指在不同物種中具有相同的基本結構和發育起源,但可能具有不同功能的器官。例如,所有哺乳動物的前肢骨骼結構基本相似,無論是用於飛行(如蝙蝠)、游泳(如鯨魚)還是奔跑(如馬)。這種相似性最好的解釋是它們源自共同的祖先形式,然後通過適應不同環境而演化成不同的形態。
痕跡器官的存在也是進化的有力證據。痕跡器官是指在某些物種中存在但已失去原有功能的結構。例如,人類的闌尾、鯨魚的髖骨殘跡,或者某些蛇類體內的殘留後肢骨骼。這些結構在現代生物中可能沒有明顯功能,但它們的存在可以通過進化來解釋:它們是祖先物種中曾經有用的器官的殘留。
胚胎學研究也為進化理論提供了重要支持。不同物種的胚胎,特別是在早期發育階段,常常表現出驚人的相似性。例如,所有脊椎動物的胚胎在早期都有魚類樣的特徵,如尾部和鰓裂。這種相似性反映了它們共同的進化歷史。著名的生物學家恩斯特·海克爾(Ernst Haeckel)曾提出"個體發生重演系統發生"的理論,雖然這個理論在細節上被證明是過於簡化的,但它捕捉到了一個重要的真理:胚胎發育過程反映了物種的進化歷史。
生物地理學為進化理論提供了另一層面的證據。物種的分布模式常常反映了它們的進化歷史和大陸漂移的影響。例如,澳大利亞獨特的動物群(如袋鼠和鴨嘴獸)可以通過它與其他大陸長期隔離的歷史來解釋。類似地,加拉帕戈斯群島上的物種與南美大陸物種的相似性,以及群島內部物種的差異,為達爾文提供了重要的進化洞見。
分子生物學的發展為進化研究提供了全新的維度。DNA序列分析使科學家能夠精確地測量不同物種之間的遺傳相似性,從而重建演化關係。這種方法不僅確認了基於形態學的許多演化關係,還揭示了一些意想不到的聯繫。例如,分子研究顯示鯨類與河馬有近親關係,這是純粹基於形態學難以發現的。此外,分子鐘技術允許科學家估算物種分化的時間,為化石記錄提供了補充。
比較基因組學研究進一步支持了進化理論。科學家發現,不同物種之間存在大量的共同基因。例如,人類和黑猩猩的基因組有超過98%的相似性。更令人驚訝的是,即使在進化上相距甚遠的物種之間也存在許多共同的基因。例如,人類和果蠅共享許多調控發育的關鍵基因。這種遺傳相似性最好的解釋是所有生物都源自共同的祖先。
直接觀察到的進化為這個理論提供了最直接的證據。雖然大多數進化過程發生在漫長的時間尺度上,但在某些情況下,我們可以直接觀察到進化的發生。細菌對抗生素的快速適應就是一個典型例子。另一個著名的例子是英國的桿斑蛾(Peppered moth),在工業革命期間,這種蛾子的顏色從淺色變為深色,以適應污染導致的環境變化。
人工選擇的結果也為自然選擇提供了有力的證據。通過有目的的育種,人類在相對短的時間內創造出了各種各樣的家畜和作物品種。例如,所有的家犬品種,從吉娃娃到大丹犬,都是通過人工選擇從狼演化而來的。這表明,如果人為選擇可以在短時間內產生如此大的變化,那麼自然選擇在漫長的地質時期內肯定能夠產生更大的變化。
實驗演化學提供了另一種直接觀察進化的方法。科學家可以在實驗室條件下對快速繁殖的生物(如細菌或果蠅)進行長期培養,觀察它們如何適應新的環境條件。這些研究不僅證實了進化的發生,還揭示了進化的速度和機制。
總的支持進化理論的證據來自多個獨立的科學領域,包括古生物學、比較解剖學、胚胎學、生物地理學、分子生物學和遺傳學等。這些證據的一致性和互補性使得進化理論成為生物學中最受支持的理論之一。重要的是,進化理論不僅解釋了已知的現象,還能夠做出可以被實驗驗證的預測。隨著科學技術的不斷進步,新的證據不斷湧現,進一步強化了進化理論的基礎。
進化理論是現代生物學的基石,但它同時也是最容易被誤解的科學理論之一。這些誤解不僅存在於公眾中,有時甚至在教育界和媒體中也會出現。這些誤解可能源於對進化理論的不完全理解、先入為主的觀念,或者是對科學方法的誤解。因此,澄清這些常見的誤解對於正確理解進化理論至關重要。
一個最常見的誤解是認為進化是"只是一個理論"。在日常用語中,"理論"常被用來表示猜測或未經證實的想法。然而,在科學中,"理論"是指一套經過嚴格測試和大量證據支持的解釋框架。進化理論就是這樣一個得到大量證據支持的科學理論,與引力理論或原子理論一樣可靠。它不僅解釋了已知的現象,還能夠做出可以被實驗驗證的預測。
另一個普遍的誤解是認為進化意味著"從猴子變成人"。這種說法嚴重誤解了進化的本質。進化理論實際上認為人類和現代猿類有共同的祖先,而不是人類從現代猿類演化而來。人類和黑猩猩等近親物種在幾百萬年前就已經分道揚鑣,各自沿著不同的進化路徑發展。因此,現代人類和現代猿類是平行進化的結果,而不是一個從另一個直接演變而來。
許多人錯誤地認為進化是一個有目的性的過程,彷彿生物在朝著某個預定的目標"進步"。這種觀點被稱為目的論(Teleology),它與科學理解的進化過程相去甚遠。實際上,進化是一個無目的的過程,它只是那些恰好適應當前環境的個體更有可能生存和繁衍的結果。進化並不必然導致更"高級"或"複雜"的生物形式,它只是產生更適應特定環境的形式。
與上述誤解相關的是認為進化總是朝著"更好"或"更完美"的方向發展。這種觀點忽視了進化的權衡性質。每種適應都有其代價,而且在一種環境中有利的特徵在另一種環境中可能是不利的。例如,人類直立行走雖然帶來了許多優勢,但也導致了背痛等問題。進化不是追求完美,而是在各種限制和權衡中尋求妥協。
一些人誤解為進化是一個純粹隨機的過程。雖然突變確實是隨機的,但自然選擇這一核心機制卻是非隨機的。自然選擇傾向於保留有利的變異,淘汰不利的變異,這導致了適應性的增加。因此,進化是隨機(突變)和非隨機(選擇)過程的結合。
另一個常見的誤解是認為"適者生存"意味著只有最強壯或最具侵略性的個體才能生存。然而,在進化意義上的"適應"(Fitness)指的是在特定環境中生存和繁衍的能力,這可能與力量或侵略性無關。例如,在某些環境中,合作或隱蔽可能比競爭更有利於生存。
很多人錯誤地認為進化是一個緩慢的過程,需要數百萬年才能看到變化。雖然大型生物的形態變化確實可能需要很長時間,但在某些情況下,進化可以非常快速。例如,細菌可以在幾周內演化出抗生素抗性。這種快速進化在小型、繁殖迅速的生物中特別明顯。
一些人認為如果進化是真的,那麼中間形式或"缺失環節"應該隨處可見。這種觀點忽視了化石形成的稀有性和不完整性。實際上,科學家已經發現了許多重要的過渡性化石,如始祖鳥(Archaeopteryx)和泰克塔利克魚(Tiktaalik)。此外,每一個生物都可以被視為其祖先和後代之間的中間形式。
另一個誤解是認為接受進化理論就必須拒絕宗教信仰。實際上,許多宗教領袖和信徒認為進化理論與他們的信仰並不矛盾。進化理論是一個科學理論,解釋自然現象,而宗教則涉及信仰和價值觀。這兩者並不必然衝突,許多科學家同時也是虔誠的宗教信徒。
有人錯誤地認為人類已經停止進化。雖然現代醫學和技術確實改變了一些選擇壓力,但這並不意味著進化已經停止。人類仍然面臨著新的選擇壓力,如抗生素耐藥性細菌的威脅。此外,文化演化在人類社會中扮演著越來越重要的角色。
一些人誤解為如果人類是通過進化而來,那麼我們就應該能夠在動物園裡看到猴子變成人。這種觀點反映了對進化時間尺度的誤解。進化是一個需要數百萬年的緩慢過程,不可能在人類的壽命範圍內直接觀察到如此巨大的變化。
另一個常見的誤解是認為進化理論無法被證實,因此不是真正的科學。這種觀點反映了對科學方法的誤解。雖然我們不能直接觀察到過去的進化過程,但我們可以通過多種證據來支持進化理論,包括化石記錄、比較解剖學、分子生物學等。此外,進化理論可以做出可被證實或證偽的預測,這是科學理論的關鍵特徵。
有人錯誤地認為接受進化理論就意味著生命沒有意義或目的。這是一個哲學問題,而不是科學問題。進化理論解釋了生命如何發展,但並不涉及生命的意義或目的。這些問題屬於哲學、倫理學和個人信仰的範疇。
最後,一個常見的誤解是認為教導進化論會導致道德淪喪或社會問題。這種觀點通常源於對"適者生存"的誤解,認為它提倡弱肉強食的社會達爾文主義。然而,進化理論是對自然現象的科學描述,不應被用來證明任何道德或社會立場。實際上,理解進化可以增進我們對生命相互聯繫的認識,促進對自然的尊重和保護。
物種形成是進化理論中的一個核心概念,它解釋了新物種如何在自然界中產生。這個過程是生物多樣性的根源,也是我們理解生命演化歷史的關鍵。物種形成通常被定義為一個物種分化為兩個或多個不同物種的過程。這個過程涉及遺傳、生態和地理等多個因素的相互作用。
在討論物種形成之前,我們首先需要了解什麼是物種。雖然物種的定義在生物學界仍有爭議,但最廣為接受的定義是生物學物種概念(Biological Species Concept)。根據這個概念,物種是一群實際或潛在可以相互交配並產生可育後代的生物個體。這個定義強調了生殖隔離在物種形成中的重要性。
物種形成的過程通常始於種群內的遺傳變異。這種變異可能源於突變、基因重組或基因流動。當種群中的不同個體擁有不同的遺傳特徵時,它們可能對環境的適應能力也不同。如果環境條件發生變化,或者種群遷移到新的環境中,這些遺傳差異可能導致某些個體比其他個體具有更高的生存和繁殖能力。
地理隔離通常被認為是物種形成的一個重要因素。當一個種群被分割成兩個或多個地理上隔離的亞群時,這些亞群可能會受到不同的選擇壓力。隨著時間的推移,這些亞群可能會逐漸積累遺傳差異,最終導致它們無法相互交配。這種過程被稱為異域物種形成(Allopatric speciation)。異域物種形成在自然界中非常常見,特別是在島嶼等相對隔離的環境中。
然而,物種形成並不總是需要完全的地理隔離。在同域物種形成(Sympatric speciation)中,新物種可以在沒有地理障礙的情況下形成。這種物種形成通常涉及生態位分化,即種群中的不同個體適應不同的生態環境。例如,某些魚類可能specializes for feeding在湖泊的不同深度,導致種群分化為兩個生態上不同的群體。隨著時間的推移,這些群體可能發展出生殖隔離機制,最終形成不同的物種。
旁域物種形成(Parapatric speciation)是介於異域和同域物種形成之間的一種形式。在這種情況下,兩個亞種群的分佈區域部分重疊,但大部分仍然是分開的。這種情況可能發生在環境梯度上,例如沿著山坡或溫度梯度。隨著時間的推移,這些部分隔離的亞群可能會演化出不同的特徵,最終形成不同的物種。
生殖隔離機制在物種形成過程中起著關鍵作用。這些機制可以分為前合子隔離機制和後合子隔離機制。前合子隔離機制防止不同種群的個體交配或阻止受精的發生。例如,不同的求偶行為、不同的開花時間或者不兼容的生殖器官結構都可以作為前合子隔離機制。後合子隔離機制則阻止雜交後代的存活或生育能力。例如,雜交後代可能無法存活,或者即使存活也可能不育。
物種形成的速度可以vary greatly。在某些情況下,物種形成可能是一個緩慢的過程,需要數百萬年的時間。然而,在其他情況下,特別是在植物中,物種形成可能發生得相對迅速。例如,通過染色體加倍(Polyploidy),新的植物物種可以在一代之內形成。這種現象在農作物中特別常見,許多重要的農作物都是多倍體。
物種形成的過程並不總是完整或不可逆的。在某些情況下,正在分化的種群可能再次融合,這個過程被稱為物種融合(Species fusion)。此外,一些物種可能保持部分的基因交流,形成所謂的物種複合體(Species complex)。這些現象提醒我們,物種形成是一個連續的過程,而不是一個非黑即白的事件。
研究物種形成的方法包括化石研究、比較基因組學、生態學觀察和實驗室實驗。化石記錄可以提供物種形成的歷史證據,顯示物種如何隨時間變化。比較基因組學允許科學家追踪物種之間的遺傳差異,並估計它們分化的時間。生態學觀察可以揭示物種如何適應不同的環境,以及這些適應如何導致種群分化。實驗室實驗,特別是使用快速繁殖的生物如果蠅或細菌,可以讓科學家直接觀察物種形成的過程。
物種形成的研究對於理解生物多樣性的起源和維持至關重要。它不僅幫助我們理解過去的進化歷史,還為預測未來的生物多樣性變化提供了基礎。例如,了解物種如何適應不同的環境條件可以幫助我們預測氣候變化對生物多樣性的影響。
物種形成的概念也有重要的實際應用。在農業中,理解作物的物種形成過程可以幫助開發新的品種。在保護生物學中,識別獨特的演化支系對於確定保護優先順序至關重要。在醫學領域,了解病原體的物種形成過程可以幫助我們預測和應對新的疾病威脅。
然而,物種形成的研究也面臨著許多挑戰。首先,物種形成通常是一個漫長的過程,難以在人類的時間尺度內直接觀察。其次,物種的定義本身就是一個複雜的問題,特別是在微生物和正在分化的種群中。此外,很多物種形成的中間階段可能已經滅絕,使得重建完整的物種形成過程變得困難。
儘管如此,隨著新技術的發展,特別是在基因組學和生物信息學領域,我們對物種形成的理解正在不斷深化。例如,全基因組測序技術使我們能夠更全面地比較不同物種的遺傳差異,從而更好地理解物種分化的遺傳基礎。同時,大數據分析技術也使我們能夠從海量的生物多樣性數據中發現物種形成的模式。
物種形成的研究也引發了一些有趣的哲學問題。例如,如果物種形成是一個漸進的過程,那麼我們如何確定一個新物種何時"誕生"?這個問題挑戰了我們對物種分類的傳統觀念,提醒我們物種可能更像是一個連續體,而不是完全分離的實體。
物種是生物分類學中的一個基本單位,也是生態學和進化生物學研究的核心概念。然而,定義"物種"這個看似簡單的問題實際上是生物學中最具爭議和複雜的問題之一。不同的生物學家可能根據他們的研究領域和研究對象採用不同的物種定義。
最廣為人知的物種定義是生物學物種概念(Biological Species Concept),由恩斯特·邁爾(Ernst Mayr)在20世紀中期提出。根據這個定義,物種是一群實際或潛在可以相互交配並產生可育後代的生物個體。這個定義強調了生殖隔離在物種形成和維持中的重要性。生物學物種概念在動物學研究中特別有用,因為它提供了一個清晰的標準來區分不同的物種。
然而,生物學物種概念也有其局限性。首先,它不適用於無性生殖的生物,如許多細菌和一些植物。其次,它難以應用於化石物種,因為我們無法確定已滅絕的生物是否能夠相互交配。此外,即使在有性生殖的生物中,也存在種間雜交的現象,這使得物種界限變得模糊。
為了解決這些問題,生物學家提出了多種替代或補充的物種概念。形態學物種概念(Morphological Species Concept)是最古老的物種定義之一,它根據生物體的形態特徵來區分物種。這個概念在古生物學和植物學中特別有用,因為它可以應用於化石和標本。然而,形態學物種概念可能會忽視隱蔽種(Cryptic species)的存在,這些物種在形態上相似但在遺傳上卻有顯著差異。
生態學物種概念(Ecological Species Concept)將物種定義為佔據相同生態位的個體群體。這個概念強調了物種在生態系統中的功能角色,但它可能難以應用於生態位相似但遺傳上distinct的物種。
系統發生學物種概念(Phylogenetic Species Concept)將物種定義為最小的可辨識的單系群(Monophyletic group)。這個概念與現代的分子系統學研究方法相容,但可能導致物種數量的大幅增加,因為它可能將地理上isolated的種群視為不同的物種。
進化物種概念(Evolutionary Species Concept)試圖結合多個物種概念的優點,將物種定義為一個單系的祖先-後代序列,該序列保持其獨特的進化角色和趨勢。這個概念在理論上很吸引人,但在實踐中可能難以應用,因為確定一個種群的"進化角色和趨勢"可能具有挑戰性。
基因庫物種概念(Genotypic Cluster Species Concept)定義物種為在基因型空間中形成distinct clusters的個體群體。這個概念利用了現代基因組學技術的優勢,可以檢測到形態上難以區分的物種差異。然而,它可能不適用於基因流動頻繁的物種群。
在微生物學領域,由於細菌和古菌主要通過無性生殖繁衍,傳統的物種概念往往難以適用。微生物學家通常使用DNA同源性或16S rRNA基因序列相似性等標準來定義物種。然而,這些標準也存在爭議,因為細菌之間頻繁的橫向基因轉移使得物種界限變得模糊。
在病毒學中,物種的定義更加複雜。病毒不符合傳統的生物定義,因為它們不能獨立複製,也不具有細胞結構。國際病毒分類委員會(International Committee on Taxonomy of Viruses, ICTV)將病毒物種定義為一個單系群的病毒,其性質使其區別於其他物種。這個定義相當靈活,允許根據不同病毒組的特性採用不同的分類標準。
在植物學中,物種的定義也面臨特殊挑戰。許多植物可以通過雜交產生可育的後代,即使它們在形態和生態上有顯著差異。此外,多倍體化(Polyploidization)在植物中很常見,可以迅速產生新的物種。因此,植物學家通常需要綜合考慮形態、生態和遺傳等多方面的證據來定義物種。
值得注意的是,物種的概念在不同的時間尺度上可能有不同的意義。在短期內,生物學物種概念可能是最實用的,因為它關注的是當前的生殖隔離。但在長期的進化尺度上,系統發生學或進化物種概念可能更有意義,因為它們反映了物種的歷史軌跡。
物種概念的多樣性反映了生物多樣性的複雜性。不同的物種概念可能導致對同一組生物體的不同分類。這種情況在分類學中被稱為分類學膨脹(Taxonomic inflation)或分類學縮減(Taxonomic deflation),取決於新的分類方法是增加還是減少了被認可的物種數量。
物種概念的爭論不僅是學術問題,還有重要的實際影響。例如,在保護生物學中,物種的定義直接影響了哪些生物群體被視為需要保護的獨立單位。在法律和政策制定中,物種的定義可能影響到哪些生物受到法律保護。
近年來,隨著基因組學技術的發展,我們對物種界限的理解變得更加複雜。全基因組測序揭示了即使在傳統上被認為是單一物種的生物群體中也存在顯著的遺傳變異。這導致了"基因組物種"概念的提出,即基於全基因組相似性來定義物種。
然而,基因組學研究也揭示了物種之間的基因流動(Gene flow)比我們之前認為的更為普遍。這種現象在植物中特別常見,但在動物中也不罕見。這挑戰了生物學物種概念的基本假設,即不同物種之間應該是生殖隔離的。
一些生物學家認為,我們應該放棄尋找一個統一的物種定義,而應該接受物種概念的多元性。他們認為,不同的研究問題和生物群體可能需要不同的物種概念。這種觀點強調了物種作為一個概念工具的靈活性,而不是將其視為自然界中客觀存在的實體。
另一方面,也有學者提出了整合不同物種概念的嘗試。例如,統一物種概念(Unified Species Concept)試圖將物種定義為進化上independent的譜系。這個概念試圖捕捉物種形成的動態過程,同時也承認物種界限的模糊性。
物種概念的討論也引發了一些哲學問題。例如,物種是否是客觀存在的自然類別,還是人為的分類概念?物種是否有本質,還是僅僅是進化連續體上的任意切割?這些問題不僅關係到我們如何理解和分類生物多樣性,也涉及到我們如何理解自然界的結構和組織。
物種形成是生物多樣性產生的核心過程,它解釋了新物種如何從現有物種中演化而來。生物學家根據物種形成過程中種群的地理分布和隔離程度,將物種形成分為幾種主要類型。每種類型都有其獨特的特徵和機制,但它們都導致了新物種的產生。
異域物種形成(Allopatric speciation)是最容易理解和最廣為接受的物種形成類型。在這種情況下,一個物種的種群被地理障礙分隔成兩個或多個隔離的亞種群。這些障礙可能是山脈、河流、海洋,甚至是人為的障礙如公路或城市。隨著時間的推移,這些隔離的亞種群可能會適應不同的環境條件,積累遺傳差異。如果這種隔離持續足夠長的時間,這些亞種群可能會演化成無法相互交配的獨立物種。
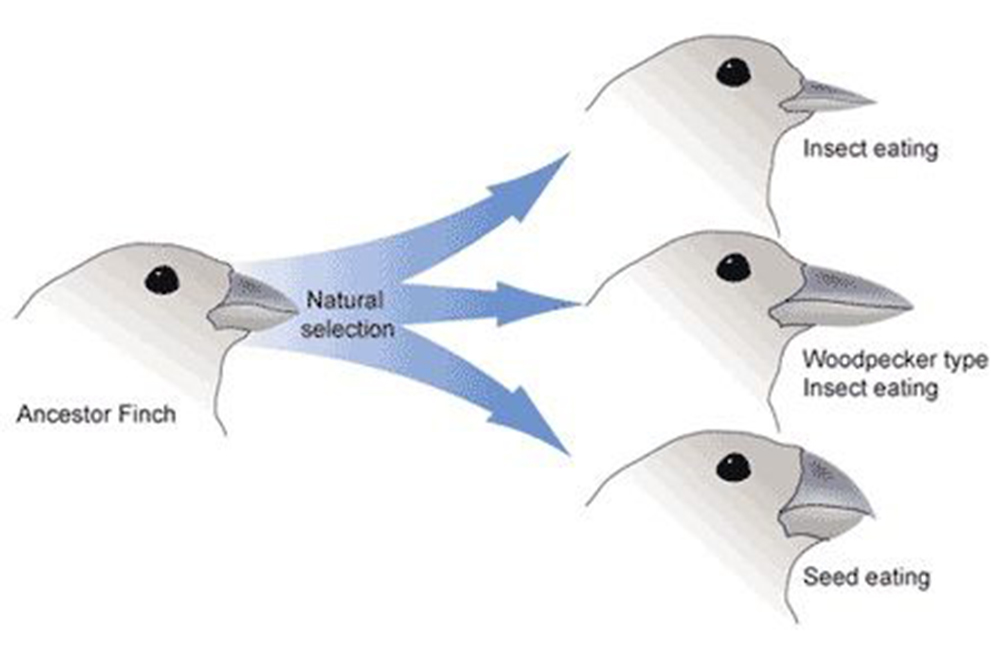
達爾文雀(也稱為加拉帕戈斯雀)的例子是異域物種形成的經典案例。這些鳥類最初來自南美大陸,但在加拉帕戈斯群島的不同島嶼上演化成了不同的物種。每個島嶼上的雀鳥適應了當地特定的食物來源,導致了喙部形狀和大小的顯著差異。這種適應最終導致了生殖隔離,形成了新的物種。
周緣物種形成(Peripatric speciation)是異域物種形成的一種特殊形式。在這種情況下,一個小的邊緣種群與主要種群隔離。由於這個小種群的基因庫有限,遺傳漂變(genetic drift)可能在物種形成過程中起到重要作用。這種物種形成方式可能解釋了一些島嶼上特有物種的起源。
同域物種形成(Sympatric speciation)是指新物種在沒有地理隔離的情況下形成。這種物種形成類型曾經被認為是非常罕見的,但近年來的研究表明它可能比previously thought要常見。同域物種形成通常涉及生態位分化,即種群中的不同個體適應不同的生態環境或資源。
同域物種形成的一個著名例子是非洲大湖區的蘆鳍科魚類(cichlid fish)。在這些湖泊中,大量的蘆鳍魚種類在相對較短的時間內演化而來,儘管它們生活在同一水體中。這些魚類適應了不同的食物來源、棲息地深度和繁殖行為,導致了reproductive isolation和新物種的形成。
旁域物種形成(Parapatric speciation)是介於異域和同域物種形成之間的一種形式。在這種情況下,兩個亞種群的分布區域部分重疊,但大部分仍然是分開的。這種情況可能發生在環境梯度上,例如沿著山坡或溫度梯度。隨著時間的推移,這些部分隔離的亞群可能會演化出不同的特徵,最終形成不同的物種。
旁域物種形成的一個例子可能是歐洲草地蜥蜴(meadow grasshopper)。這種蜥蜴在不同海拔高度上顯示出不同的染色體結構,但它們的分布區域是連續的,沒有明顯的地理障礙。
多倍體物種形成(Polyploid speciation)是植物中特別常見的一種快速物種形成方式。多倍體是指生物體擁有三套或更多套染色體,而不是通常的兩套。多倍體可能源於同一物種內的染色體加倍(自多倍體,autopolyploidy),或者源於不同物種之間的雜交後染色體加倍(異多倍體,allopolyploidy)。
多倍體物種形成的一個著名例子是普通小麥(Triticum aestivum)的起源。普通小麥是一個六倍體,它是通過兩次異多倍體事件形成的。首先,兩個二倍體物種雜交形成了四倍體小麥,然後這個四倍體小麥又與另一個二倍體物種雜交,最終形成了六倍體的普通小麥。
雜交物種形成(Hybrid speciation)是指通過兩個不同物種雜交而產生新物種的過程。雖然大多數種間雜交會產生不育的後代,但在某些情況下,雜交可能導致新物種的形成。這在植物中特別常見,尤其是當雜交伴隨著多倍體化時。
向日葵屬(Helianthus)中的三個物種就是雜交物種形成的例子。Helianthus anomalus、H. deserticola 和 H. paradoxus 都是通過兩個祖先物種 H. annuus 和 H. petiolaris 的雜交而形成的。這些新物種適應了與其親本物種不同的生態環境。
量子物種形成(Quantum speciation)是指通過突然的大規模遺傳變化而快速形成新物種的過程。這種物種形成可能涉及染色體重排或其他重大的遺傳改變。雖然這種物種形成方式可能不如其他類型常見,但它可能解釋了某些物種的急速演化。
生態物種形成(Ecological speciation)強調自然選擇在物種形成中的作用。在這種情況下,適應不同生態位的選擇壓力導致了種群的分化和最終的生殖隔離。這種物種形成可能發生在任何地理情況下,無論是異域、同域還是旁域。
重要的是要注意,這些物種形成類型並不是互斥的,實際的物種形成過程可能涉及多種機制。例如,最初由地理隔離引起的種群分化(異域過程)可能後來通過生態適應(生態物種形成)而加強。
此外,物種形成的速度可能因不同的類型而異。多倍體物種形成可能在一代之內發生,而異域物種形成可能需要數百萬年的時間。然而,即使是被認為緩慢的物種形成過程,在地質時間尺度上也可能相對迅速。
隨著基因組學技術的發展,我們對物種形成過程的理解正在不斷深化。全基因組測序使我們能夠識別參與適應和生殖隔離的特定基因,並追踪物種形成過程中的基因流動模式。這些新技術正在揭示物種形成的複雜性和多樣性,挑戰了我們對這一過程的傳統理解。
物種重新聯繫是進化生物學中一個引人入勝的主題,它探討了曾經分離的物種或種群如何再次相遇並互動的過程。這個概念與物種形成密切相關,但關注的是物種分化後的命運。當曾經分離的種群或物種重新接觸時,可能會發生各種有趣的進化現象,包括雜交、基因滲透、競爭、或進一步的分化。
在自然界中,物種重新聯繫可能由多種因素引起。地質事件,如大陸漂移、山脈形成或消失、海平面變化等,都可能導致曾經分離的種群重新接觸。氣候變化也可能引起物種分布範圍的擴大或縮小,導致原本分離的物種重新相遇。人類活動,如物種引入或棲息地改變,也越來越成為物種重新聯繫的重要因素。
當兩個分離的種群重新接觸時,可能會出現幾種不同的結果。一種可能是兩個種群已經演化出足夠的差異,無法再進行有效的基因交流,它們可能會作為獨立的物種共存,或者一個可能會排擠另一個。另一種可能是兩個種群仍然能夠雜交,這可能導致基因滲透,甚至可能導致兩個種群的融合。
雜交區(Hybrid zone)是研究物種重新聯繫的重要場所。雜交區是兩個相近物種或亞種的分布區重疊並產生雜交後代的區域。這些區域為研究物種間的基因流動、適應性和生殖隔離機制提供了寶貴的自然實驗室。著名的雜交區例子包括歐洲的褐蠟嘴雀(Certhia familiaris)和短趾蠟嘴雀(C. brachydactyla)之間的雜交區,以及北美的藍翅和金翅林鶯之間的雜交區。
基因滲透(Introgression)是物種重新聯繫過程中常見的現象。它指的是通過雜交和回交,一個物種的基因轉移到另一個物種的基因庫中的過程。基因滲透可能導致適應性狀的轉移,增加受體物種的遺傳多樣性,有時甚至可能導致適應性輻射。然而,過度的基因滲透也可能威脅物種的完整性,在某些情況下導致物種的融合或滅絕。
人類活動加速了物種重新聯繫的過程。隨著全球化的發展,許多物種被有意或無意地引入到新的環境中。這些入侵物種可能與本地近親物種接觸,導致複雜的進化互動。例如,在北美,引入的歐洲綠蟋蟀(Orthoptera)與本地物種雜交,產生了具有新的行為和生態特徵的混合種群。
氣候變化也正在改變物種的分布範圍,導致原本分離的物種重新接觸。隨著全球變暖,許多物種正在向極地或高海拔地區遷移,這可能導致原本分離的近緣物種相遇。例如,在北美,隨著氣候變暖,南方的紅樫(Quercus rubra)正在向北擴展,與北方的黑櫟(Q. velutina)的分布區重疊,可能導致兩個物種之間的雜交增加。
物種重新聯繫可能對生態系統產生深遠的影響。當曾經分離的物種重新接觸時,它們可能競爭相同的資源,改變當地的生態平衡。在某些情況下,一個物種可能完全取代另一個物種。例如,在北美的五大湖地區,入侵的歐洲丁香(Syringa vulgaris)正在與本地的美洲丁香(Syringa americana)競爭,並在某些地區取代了後者。
從進化的角度來看,物種重新聯繫可能產生新的選擇壓力。例如,為了避免有害的雜交,物種可能演化出更強的生殖隔離機制。這種現象被稱為增強(Reinforcement),它可能加速物種的分化過程。另一方面,如果雜交產生的後代具有適應優勢,可能會促進適應性狀的快速進化或新物種的形成。
物種重新聯繫還為我們提供了研究物種形成過程的機會。通過研究不同程度分化的種群如何相互作用,我們可以更好地理解生殖隔離機制是如何演化的。例如,研究者發現,在雜交區中,生殖隔離的強度往往與種群分離的時間成正比,這支持了物種形成是一個漸進過程的觀點。
在保護生物學中,理解物種重新聯繫的過程至關重要。當瀕危物種的種群被隔離時,保護生物學家可能考慮重新連接這些種群以增加遺傳多樣性。然而,這種做法也可能帶來風險,如外洩抑制(Outbreeding depression),即由於局部適應的喪失導致混合後代適應度下降。因此,在實施此類保護措施時需要仔細權衡利弊。
分子生物學技術的發展為研究物種重新聯繫提供了新的工具。基因組學方法使我們能夠精確地量化不同種群之間的基因流動,識別參與適應和生殖隔離的基因,以及追踪雜交事件的歷史。例如,通過全基因組測序,研究者發現現代人類基因組中含有少量尼安德特人和丹尼索瓦人的DNA,這揭示了早期人類種群之間的複雜互動歷史。
物種重新聯繫的研究也引發了一些有趣的進化問題。例如,如果兩個分離的種群在隔離期間積累了互補的有害突變,那麼它們重新接觸後的雜交可能會產生比父母更適應的後代。這種現象被稱為雜種優勢(Hybrid vigor)或異質雜種(Heterosis)。另一方面,如果兩個種群適應了不同的環境,它們的雜交後代可能在任何環境中都表現不佳,這種現象被稱為雜種崩潰(Hybrid breakdown)。
物種重新聯繫的研究還涉及了一些哲學問題。例如,如果兩個曾經分離的種群重新融合,我們是否應該將它們視為一個物種?這個問題挑戰了我們對物種概念的理解,提醒我們物種可能是一個動態的、而非靜態的概念。
在應用層面,理解物種重新聯繫的過程對農業和醫學也有重要意義。在農業中,不同作物品種或農作物與其野生親緣種之間的雜交可能導致新的適應性狀的產生,但也可能帶來基因汙染的風險。在醫學領域,了解病原體種群之間的重新聯繫和基因交流對於預測和控制疾病的傳播至關重要。
總的物種重新聯繫是一個複雜的生物學過程,涉及進化、生態、遺傳學等多個領域。它不僅幫助我們理解生物多樣性的動態變化,也為我們應對全球變化帶來的挑戰提供了重要的見解。隨著研究方法的不斷進步,我們對這一過程的理解也在不斷深化,揭示了生命演化的複雜性和多樣性。
物種形成的速率是進化生物學中一個重要而複雜的主題。它探討了新物種產生的頻率和速度,這對於理解生物多樣性的起源和維持至關重要。物種形成的速率可能受到多種因素的影響,包括環境條件、生物的特徵、地質事件等。研究物種形成的速率不僅有助於我們理解過去的進化歷史,還能幫助我們預測未來的生物多樣性變化。
物種形成的速率在不同的生物群體和不同的時期可能有很大差異。有些物種可能在幾百萬年內保持相對穩定,而其他物種可能在相對較短的時間內經歷快速的分化。例如,非洲大湖區的蘆鳍科魚類(Cichlid fish)就展示了極快的物種形成速率,在幾十萬年內產生了數百個新物種。相比之下,被稱為"活化石"的物種,如鱟(Horseshoe crab)和銀杏(Ginkgo biloba),在數億年的時間裡幾乎沒有發生顯著變化。
研究物種形成速率的一個主要方法是分析化石記錄。通過研究不同地質時期的化石分布,科學家可以估算物種出現和滅絕的速率。然而,這種方法也面臨著一些挑戰。首先,化石記錄通常是不完整的,特別是對於軟體生物或生活在不利於化石形成的環境中的生物。其次,確定化石代表的是新物種還是已知物種的變異有時可能很困難。
分子鐘方法為估算物種形成速率提供了另一種途徑。這種方法基於DNA或蛋白質序列隨時間積累變異的假設。通過比較不同物種的基因序列並結合化石證據進行校準,科學家可以估算物種分化的時間。然而,分子鐘方法也有其局限性。不同基因和不同生物的進化速率可能不同,這可能導致估算的誤差。
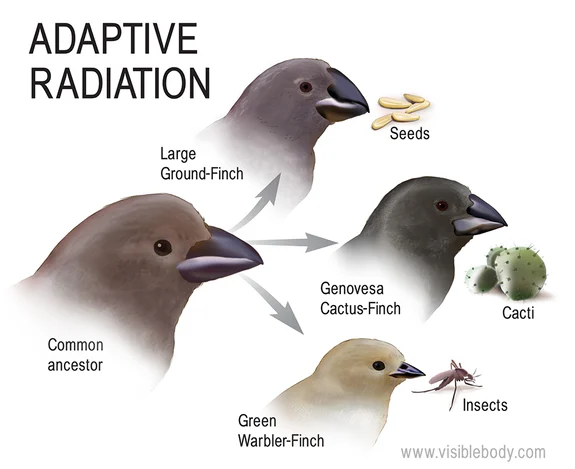
物種形成的速率可能受到多種因素的影響。環境變化通常被認為是促進物種形成的重要因素。當環境發生顯著變化時,可能會產生新的生態位,為適應性輻射提供機會。例如,在新的島嶼形成後,移居到島上的生物可能會快速演化以適應新的環境,產生一系列新的物種。這種現象在夏威夷群島和加拉帕戈斯群島上尤為明顯。
生物的特徵也可能影響物種形成的速率。例如,具有較短世代時間的生物(如細菌和某些昆蟲)可能比長壽的生物(如大型哺乳動物)有更快的物種形成速率。這是因為短世代時間允許遺傳變異更快地積累和傳播。同樣,具有較強散布能力的生物可能更容易colonize新的環境,從而增加物種形成的機會。
地質事件也可能顯著影響物種形成的速率。大規模的地質變化,如大陸漂移、山脈形成或冰川期,可能導致生物群體的分離或新棲息地的形成,從而促進物種形成。例如,安第斯山脈的隆起被認為是南美洲生物多樣性豐富的重要原因之一,它創造了多樣化的環境並分隔了許多生物群體。
競爭和捕食關係也可能影響物種形成的速率。根據範布倫敦河紅皇后假說(Van Valen's Red Queen hypothesis),物種需要不斷進化以維持其在生態系統中的位置。這種持續的進化壓力可能導致加速的物種形成。例如,植物和授粉者之間的協同進化可能導致兩個群體的快速分化。
物種形成的速率可能不是恆定的,而是隨時間波動。古生物學證據表明,地球歷史上存在幾次生物多樣性的快速增加期,被稱為演化輻射(Evolutionary radiation)。這些期間的物種形成速率遠高於平均水準。例如,寒武紀大爆發(Cambrian explosion)期間,幾乎所有現代動物門類在相對較短的時間內出現。
相反,也存在物種形成速率明顯降低的時期。這通常與大規模滅絕事件有關。在這些事件之後,生物多樣性的恢復可能需要數百萬年的時間。例如,在二疊紀末期的大滅絕之後,三疊紀早期的物種形成速率相對較低,直到生態系統逐漸恢復後才開始加速。
近年來,全基因組測序技術的發展為研究物種形成速率提供了新的工具。通過比較近緣物種的全基因組,科學家可以更精確地估算物種分化的時間,並識別可能參與物種形成過程的基因。這種方法還揭示了物種形成可能比我們之前認為的更為複雜,涉及基因組的不同區域以不同的速率演化。
人類活動也正在以前所未有的方式影響物種形成的速率。一方面,人類導致的棲息地破壞和氣候變化正在加速許多物種的滅絕,可能減緩整體的物種形成速率。另一方面,人類活動也創造了新的選擇壓力和生態位,可能在某些情況下加速物種形成。例如,在城市環境中,一些動植物正在快速適應新的條件,可能導致新的城市適應型的出現。
理解物種形成的速率對於保護生物學具有重要意義。如果我們知道物種通常需要多長時間才能形成,我們就能更好地評估生態系統恢復的時間尺度,並制定更有效的保護策略。例如,如果某個生態系統的物種形成速率非常慢,那麼保護現有物種就變得尤為重要,因為一旦物種滅絕,可能需要很長時間才能通過自然過程恢復多樣性。
物種形成速率的研究還涉及一些有趣的理論問題。例如,是否存在物種形成速率的上限?生態系統是否有承載新物種的最大容量?這些問題不僅關係到我們對進化過程的理解,還涉及生態系統的穩定性和resilience。
在實際應用中,理解物種形成的速率對農業和醫療也有重要影響。在農業中,了解作物和害蟲的進化速率可以幫助我們開發更持久的抗性品種和更有效的害蟲管理策略。在醫療領域,理解病原體的進化速率對於開發疫苗和控制傳染病至關重要。
總的物種形成的速率是一個複雜的主題,涉及多個生物學分支的知識。它不僅幫助我們理解生物多樣性的歷史和現狀,還為預測和管理未來的生物多樣性變化提供了重要的視角。隨著研究方法的不斷進步,我們對物種形成速率的理解也在不斷深化,揭示了生命演化的複雜性和動態性。
群體進化是進化生物學的核心概念之一,它研究的是遺傳變異在群體中隨時間變化的過程。這個領域結合了遺傳學、生態學和統計學的原理,以理解和預測群體中基因頻率的變化。群體進化的研究不僅幫助我們理解生物多樣性的起源和維持,還為醫學、農業和保護生物學等應用領域提供了重要的理論基礎。
群體進化的基本單位是群體,而不是個體。在這個背景下,群體被定義為在特定時間和空間內能夠相互交配的同種個體的集合。群體可大可小,從幾個個體到數百萬個體不等。重要的是,群體內的個體共享一個共同的基因庫,這個基因庫是群體進化的基礎。
遺傳變異是群體進化的原材料。沒有遺傳變異,進化就無法發生。在一個群體中,幾乎每個基因都可能有多個變體,稱為等位基因(Alleles)。這些等位基因在群體中的相對頻率可以隨時間而變化,這種變化就是群體進化的本質。
影響群體進化的主要因素包括突變、自然選擇、遺傳漂變、基因流動和非隨機交配。突變是新遺傳變異的最終來源。雖然大多數突變可能是有害的或中性的,但偶爾也會產生有利的突變,這些突變可能會在群體中擴散。
自然選擇是達爾文進化理論的核心機制。它基於三個關鍵觀察:群體中存在遺傳變異,這些變異部分是可遺傳的,而且一些變異可能影響個體的生存和繁殖能力。那些具有有利特徵的個體更有可能存活和繁衍,從而將其基因傳給下一代。隨著時間的推移,這可能導致有利等位基因在群體中變得更加普遍。
遺傳漂變(Genetic drift)是指由於機會因素導致的等位基因頻率的隨機變化。這種效應在小群體中特別明顯,因為小樣本更容易受到隨機波動的影響。遺傳漂變可能導致某些等位基因在群體中固定或消失,即使這些等位基因對適應度沒有影響。
基因流動是指由於個體或配子的移動而導致的基因在不同群體之間的交換。基因流動可以引入新的遺傳變異,從而增加群體的遺傳多樣性。然而,過多的基因流動也可能阻礙局部適應,因為它可能使得適應特定環境的等位基因難以在群體中累積。
非隨機交配,如近親交配或正向同配交配,也可能影響群體的遺傳結構。近親交配可能增加純合子的比例,而降低雜合子的比例。這可能導致有害隱性等位基因表現出來,造成近交衰退(Inbreeding depression)。
哈迪-溫伯格平衡(Hardy-Weinberg equilibrium)是理解群體進化的重要概念。它描述了在理想條件下(大群體、隨機交配、無選擇、無突變、無遺傳漂變、無基因流動),等位基因和基因型頻率將保持不變的情況。雖然自然群體很少達到這種平衡狀態,但哈迪-溫伯格原理提供了一個重要的理論基準,用來衡量實際群體的偏離程度。
適應度(Fitness)是群體進化研究中的另一個關鍵概念。在進化意義上,適應度指的是一個個體將其基因傳遞給下一代的相對能力。適應度不僅取決於生存能力,還取決於繁殖成功。高適應度的個體更有可能將其基因傳給後代,從而影響群體的遺傳組成。
群體進化的研究方法包括野外觀察、實驗室實驗和理論模型。野外研究可以提供群體在自然條件下如何演化的直接證據。例如,對達爾文雀喙部大小的長期研究揭示了自然選擇如何影響這些特徵的進化。實驗室研究,特別是使用微生物或果蠅等短生命週期生物的實驗,可以讓我們直接觀察進化過程。理論模型,如數學模型和計算機模擬,則幫助我們理解複雜的進化過程。
群體基因組學的發展為群體進化研究提供了新的工具。全基因組測序技術使我們能夠比較同一群體中不同個體的全基因組,或比較不同群體之間的基因組差異。這種方法可以揭示選擇作用的基因組區域,追踪基因流動的模式,並估算群體的有效大小。
群體進化的研究對多個應用領域有重要影響。在醫學方面,理解病原體群體的進化可以幫助我們預測和控制傳染病的傳播,以及理解抗藥性的產生。在農業中,群體進化的原理被用於作物和牲畜的育種計劃,以及害蟲管理策略的制定。在保護生物學中,了解瀕危物種群體的遺傳結構對於制定有效的保護策略至關重要。
群體進化的研究還揭示了一些有趣的現象。例如,頻率依賴選擇(Frequency-dependent selection)是指一個等位基因的適應度取決於它在群體中的頻率。這種選擇可能導致多態性的維持。另一個例子是性選擇(Sexual selection),它解釋了為什麼某些看似不利於生存的特徵(如孔雀的華麗尾羽)能夠進化。
平衡選擇(Balancing selection)是另一個重要的群體進化概念。它解釋了為什麼某些遺傳變異能在群體中長期維持。例如,鐮狀細胞性狀在瘧疾流行區的人群中維持高頻率,就是因為雜合子對瘧疾具有抵抗力。
群體進化的研究也面臨一些挑戰。例如,很多重要的進化過程發生在長時間尺度上,難以直接觀察。此外,自然群體常常很大且分布廣泛,難以全面採樣。環境的複雜性和變化性也使得解釋觀察到的遺傳變化變得困難。
最近的研究強調了表觀遺傳變異在群體進化中的作用。表觀遺傳修飾,如DNA甲基化,可以影響基因表達而不改變DNA序列。這些修飾有時可以跨代傳遞,為群體適應環境變化提供了一種快速的機制。
群體進化的研究還涉及一些有趣的理論問題。例如,群體如何在保持足夠的遺傳變異以適應未來變化和優化當前適應之間取得平衡?自然選擇是否總是導致最優解,或者它是否常常被歷史偶然性和發展限制所束縛?
總的群體進化是一個複雜而動態的過程,涉及多種力量的相互作用。理解這個過程不僅幫助我們解釋生物多樣性的起源和維持,還為我們應對諸如疾病控制、作物改良和生物多樣性保護等實際問題提供了重要的理論基礎。隨著研究方法的不斷進步,特別是基因組學技術的發展,我們對群體進化的理解正在不斷深化,揭示了生命演化的複雜性和多樣性。
群體遺傳學是研究群體中基因頻率變化的科學,它結合了孟德爾遺傳學、達爾文進化論和數學統計學的原理。這個領域主要關注種群內基因型和表型頻率的變化,以及影響這些變化的進化力量。
群體遺傳學的核心概念是基因庫(gene pool),指的是一個群體中所有等位基因的總和。研究人員通過觀察基因庫中等位基因頻率的變化來了解群體的進化過程。影響等位基因頻率的主要因素包括自然選擇、基因漂變、基因流動和突變。
自然選擇是達爾文進化論的核心機制,它根據個體的適應度(fitness)來影響基因頻率。適應度較高的個體更有可能存活和繁衍後代,從而將其基因傳遞給下一代。隨著時間的推移,有利的等位基因在群體中的頻率會增加。例如,在工業革命期間,英國的樺尺蛾(peppered moth)經歷了顏色從淺色到深色的演變,這就是自然選擇的典型案例。
基因漂變(genetic drift)是指由於隨機抽樣誤差導致的等位基因頻率變化。這種效應在小群體中特別明顯,因為小樣本更容易受到隨機波動的影響。基因漂變可能導致某些等位基因在群體中完全消失,或者固定下來(頻率達到100%)。奠基者效應(founder effect)和瓶頸效應(bottleneck effect)是兩種特殊的基因漂變形式,它們都會導致群體遺傳多樣性的減少。
基因流動(gene flow)指的是不同群體之間的基因交流,通常通過個體的遷移或花粉的傳播實現。基因流動可以引入新的等位基因,增加群體的遺傳多樣性,同時也可能削弱群體間的遺傳差異。在人類群體中,由於現代交通和全球化,基因流動變得越來越普遍。
突變是產生新等位基因的根本來源。雖然大多數突變是有害的或中性的,但偶爾也會出現有益突變,為自然選擇提供原材料。突變率通常很低,但在某些情況下(如暴露於輻射或某些化學物質)可能會增加。
哈迪-溫伯格平衡(Hardy-Weinberg equilibrium)是群體遺傳學的基本原理之一。它描述了在理想條件下(大群體、隨機交配、無選擇壓力、無突變、無遷移),等位基因和基因型頻率在世代間保持不變的情況。這個原理為研究實際群體中的偏離提供了一個基準。
群體遺傳學的研究方法包括數學模型、統計分析和實驗研究。數學模型如擴散方程(diffusion equation)可以描述等位基因頻率隨時間的變化。統計方法如F統計量(F-statistics)用於量化群體間的遺傳分化程度。實驗研究則可以直接觀察生物體在不同選擇壓力下的進化過程。
在分子水準上,群體遺傳學利用DNA序列數據來研究遺傳變異。中性理論(neutral theory)假設大多數分子水準的進化變化是由中性突變和基因漂變驅動的,而不是自然選擇。連鎖不平衡(linkage disequilibrium)分析可以揭示基因間的相互作用和選擇的痕跡。
群體遺傳學在許多領域都有重要應用。在保護生物學中,它幫助評估瀕危物種的遺傳多樣性和制定保護策略。在醫學遺傳學中,群體遺傳學的原理用於研究疾病相關基因的分布和頻率。在農業中,它指導作物和畜禽的育種計劃。在法醫學中,群體遺傳學的知識用於解釋DNA證據。
總的來說,群體遺傳學為我們理解生物進化和適應提供了強大的理論框架和工具。它不僅解釋了生物多樣性的形成過程,也為預測未來的進化趨勢提供了基礎。隨著基因組學技術的進步,群體遺傳學正在進入一個新的發展階段,能夠更全面地描述和分析複雜生物系統中的遺傳變異。
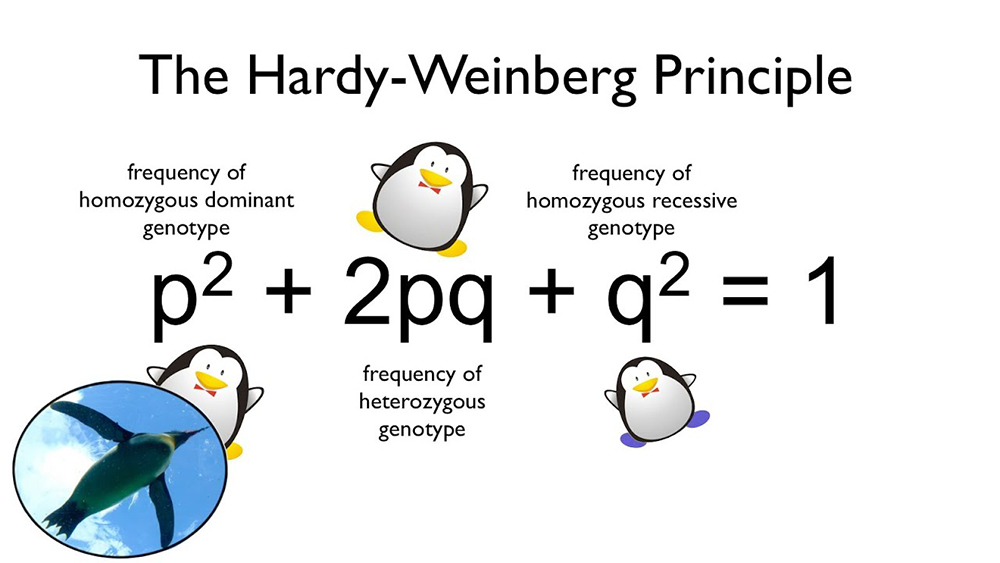
哈迪-溫伯格平衡原理是群體遺傳學中的一個基本概念,它由英國數學家G.H.哈迪和德國醫生威廉·溫伯格於1908年獨立提出。這個原理描述了在理想條件下,一個大型群體中的等位基因頻率和基因型頻率在世代之間保持不變的狀態。這個原理為理解和預測群體中基因頻率的變化提供了一個重要的理論基礎。
哈迪-溫伯格平衡原理的核心假設包括:群體規模足夠大、隨機交配、沒有突變、沒有選擇壓力、沒有遷移(即沒有基因流動)、以及世代不重疊。在這些條件下,如果一個群體達到平衡狀態,那麼它的基因型頻率將保持不變,除非有外部因素干擾。
讓我們以一個簡單的例子來說明這個原理。假設在一個群體中,有一個基因座位有兩個等位基因A和a。我們用p表示A的頻率,q表示a的頻率。根據等位基因頻率相加等於1的原則,我們有p + q = 1。在隨機交配的條件下,下一代中各種基因型的頻率可以用這些等位基因頻率的平方和積來表示:
AA的頻率 = p² Aa的頻率 = 2pq aa的頻率 = q²
這就是著名的哈迪-溫伯格方程:p² + 2pq + q² = 1。這個等式表明,在理想條件下,群體中的基因型頻率將保持穩定不變。
哈迪-溫伯格平衡原理的一個重要應用是它提供了一個理論基準,用來檢測實際群體中是否存在進化力量的作用。如果一個群體的基因型頻率偏離了哈迪-溫伯格平衡,那麼我們可以推斷可能有某些因素(如選擇、突變、遷移等)在影響這個群體的遺傳組成。
然而,需要注意的是,實際的生物群體很少能完全符合哈迪-溫伯格平衡的所有假設。例如,自然群體的大小通常是有限的,這可能導致基因漂變的發生。此外,自然選擇在許多情況下都在起作用,某些基因型可能具有生存或繁殖優勢。突變雖然罕見,但確實會不斷產生新的等位基因。遷移或基因流動在許多群體中也是常見的現象。因此,哈迪-溫伯格平衡更多地被視為一個理論模型,而不是對自然群體的精確描述。
儘管如此,哈迪-溫伯格平衡原理仍然是群體遺傳學研究的重要工具。它可以用來估算隱性等位基因的頻率。例如,如果一個隱性遺傳病在群體中的發病率為q²,那麼我們可以計算出致病等位基因的頻率q,以及攜帶者(雜合子)的頻率2pq。這對於理解遺傳病的傳播和預防有重要意義。
在醫學遺傳學中,哈迪-溫伯格平衡原理被廣泛用於研究疾病相關基因的分布。通過比較患病群體和健康群體的基因型頻率,研究人員可以識別出與疾病風險相關的基因變異。這種方法被稱為基因關聯研究(genetic association studies),它為許多複雜疾病的遺傳基礎研究做出了重要貢獻。
在保護生物學中,哈迪-溫伯格平衡原理可以幫助評估瀕危物種的遺傳健康狀況。如果一個小群體長期偏離哈迪-溫伯格平衡,可能表明它正在經歷嚴重的基因漂變或近親繁殖,這可能威脅到該物種的長期生存。
哈迪-溫伯格平衡原理也在法醫學中發揮重要作用。在分析DNA證據時,需要知道特定基因標記在一般群體中的頻率。通過應用哈迪-溫伯格原理,法醫科學家可以從有限的樣本數據中推斷出更廣泛群體的基因型頻率。
在農業和畜牧業中,哈迪-溫伯格平衡原理被用於設計育種策略。育種者可以利用這個原理來預測特定交配方案可能產生的後代基因型分布,從而選擇最有效的育種策略來獲得所需的性狀。
值得注意的是,哈迪-溫伯格平衡原理最初是為單基因性狀(monogenic traits)提出的,但它的原理可以擴展到多基因性狀(polygenic traits)。對於涉及多個基因的複雜性狀,我們可以對每個基因座位單獨應用哈迪-溫伯格原理,前提是這些基因座位之間沒有連鎖不平衡(linkage disequilibrium)。
哈迪-溫伯格平衡原理還為理解進化過程提供了洞見。它表明,在沒有外部力量作用的情況下,單純的孟德爾遺傳不會改變群體的遺傳組成。這一發現解決了達爾文進化論面臨的一個重要難題:如何解釋有利變異在群體中的保留和擴散。哈迪-溫伯格平衡原理表明,一旦有利變異出現在群體中,它不會因為與其他等位基因的混合而"稀釋"掉,而是可以在群體中穩定存在,為自然選擇提供原材料。
然而,哈迪-溫伯格平衡原理也有其局限性。它假設基因是獨立遺傳的,但實際上許多基因是連鎖的,即它們位於同一條染色體上,傾向於一起遺傳。連鎖不平衡可能導致基因型頻率偏離哈迪-溫伯格預期。此外,該原理也沒有考慮表觀遺傳(epigenetic)效應,這些效應可能影響基因的表達而不改變DNA序列。
在教學和研究中,哈迪-溫伯格平衡原理常常被用作引入更複雜群體遺傳學概念的起點。通過研究實際群體偏離平衡的方式和程度,研究人員可以推斷出影響群體進化的具體因素。例如,如果觀察到的純合子頻率高於預期,可能表明存在同型配子偏好(assortative mating)或近親繁殖。如果某些基因型的頻率低於預期,可能暗示這些基因型受到負向選擇。
總之,哈迪-溫伯格平衡原理是群體遺傳學的基石之一。它不僅提供了理解群體遺傳變異的理論框架,還為研究進化過程、疾病遺傳學和生物多樣性保護等領域提供了重要工具。儘管實際生物群體很少能完全符合其假設,但這個原理仍然是分析和解釋群體遺傳數據的重要基準。隨著分子生物學技術的進步和大數據分析方法的發展,哈迪-溫伯格平衡原理在現代生物學研究中繼續發揮著重要作用。
遺傳變異與漂變是群體遺傳學中的兩個核心概念,它們共同塑造了生物群體的遺傳結構和進化軌跡。遺傳變異指的是群體中個體之間在基因組成上的差異,而遺傳漂變則是由於隨機抽樣誤差導致的等位基因頻率變化。這兩個過程相互作用,影響著物種的適應性和長期生存。
遺傳變異是進化的原材料。沒有遺傳變異,自然選擇就無法發揮作用,物種也就無法適應環境變化。遺傳變異的來源主要包括突變、重組和基因流動。突變是DNA序列的改變,可能由於複製錯誤、環境因素(如輻射或化學物質)或病毒感染等原因引起。大多數突變是中性的,不會對生物體產生明顯影響;一些突變是有害的,可能降低個體的適應度;極少數突變是有益的,可能增強個體在特定環境中的生存和繁殖能力。
重組是在有性生殖過程中,親本基因的新組合。減數分裂時的交叉互換(crossing over)和配子的隨機結合都會產生新的基因組合。重組可以將有利的等位基因組合在一起,也可能打破不利的基因組合,從而增加群體的適應潛力。基因流動則是指不同群體之間的基因交流,通常通過個體遷移或花粉傳播實現。基因流動可以引入新的遺傳變異,增加群體的遺傳多樣性。
遺傳變異可以在多個層次上觀察到。在核苷酸水準上,單核苷酸多態性(SNP, Single Nucleotide Polymorphism)是最常見的變異形式。在基因水準上,等位基因的變異可能導致蛋白質序列或表達水準的差異。在染色體水準上,結構變異如缺失、重複、倒位和易位也是重要的變異來源。在基因組水準上,整個染色體數目的變化(如多倍體)可能導致顯著的表型差異。
測量遺傳變異的方法有多種。在分子水準上,可以通過DNA測序或基因分型技術直接觀察核苷酸或等位基因的變異。在群體水準上,常用的指標包括等位基因多樣性(每個基因座的平均等位基因數)、雜合度(雜合子在群體中的比例)和核苷酸多樣性(隨機選擇的兩個序列之間的平均差異)。這些指標可以反映群體的遺傳健康狀況和進化潛力。
遺傳漂變是另一個影響群體遺傳結構的重要因素。它指的是由於隨機抽樣誤差導致的等位基因頻率變化。在每一代,不是所有的個體都能成功繁衍後代,而是只有一部分個體的基因被傳遞到下一代。這個過程可以看作是從群體的基因庫中進行隨機抽樣。在大群體中,這種隨機波動通常很小,但在小群體中,遺傳漂變可能導致等位基因頻率的顯著變化,甚至導致某些等位基因的丟失或固定。
遺傳漂變的強度與群體的有效大小(effective population size)成反比。有效群體大小是指能夠對下一代的基因組成產生影響的繁殖個體數量。它通常小於實際的群體大小,因為並非所有個體都能平等地貢獻其基因到下一代。例如,如果一個群體中只有少數個體壟斷了繁殖機會,那麼即使總體數量很大,其有效群體大小也會很小。
遺傳漂變可能導致兩個重要的遺傳現象:奠基者效應(founder effect)和瓶頸效應(bottleneck effect)。奠基者效應發生在一小群個體離開原群體並建立新群體時。由於樣本量小,新群體的基因庫可能與原群體有很大差異。瓶頸效應則是指群體規模急劇減少後,遺傳多樣性顯著降低的現象。這兩種效應都可能導致稀有等位基因的丟失和某些等位基因頻率的顯著變化。
遺傳漂變的一個重要特徵是它是無方向性的。與自然選擇不同,遺傳漂變不會偏好有利或不利的等位基因,而是隨機地改變所有等位基因的頻率。長期來看,遺傳漂變傾向於減少群體的遺傳變異,因為它最終會導致等位基因的丟失或固定。這種效應在小群體中特別明顯,可能導致遺傳多樣性的顯著降低。
遺傳漂變的影響可以通過多種方法來研究。在實驗室條件下,可以通過控制繁殖群體的大小來觀察遺傳漂變的效應。在自然群體中,可以通過比較不同大小群體的遺傳多樣性來推斷遺傳漂變的作用。此外,分子標記如微衛星DNA(microsatellite DNA)和單核苷酸多態性可以用來追蹤等位基因頻率的變化。
遺傳變異和遺傳漂變的相互作用對物種的進化有深遠影響。一方面,遺傳變異為自然選擇提供了原材料,使物種能夠適應環境變化。另一方面,遺傳漂變可能導致有利變異的丟失,特別是在小群體中。這種相互作用在不同的情況下可能產生不同的結果。在大群體中,選擇可能是主導力量,而在小群體中,遺傳漂變可能超過選擇的作用。
在保護生物學中,理解遺傳變異和漂變的重要性不言而喻。對於瀕危物種,維持足夠的遺傳變異對其長期生存至關重要。保護策略可能包括增加群體大小、建立遷移走廊以促進基因流動,或者在極端情況下進行人工基因交流。同時,認識到遺傳漂變的作用可以幫助解釋小群體中觀察到的異常基因頻率,並指導制定適當的管理措施。
在人類群體中,遺傳變異和漂變也扮演著重要角色。人類群體的遺傳結構受到歷史遷徙、人口波動和文化因素的影響。例如,某些人群中特定遺傳疾病的高發可能是由於奠基者效應或遺傳漂變,而不是選擇壓力的結果。理解這些過程對於醫學遺傳學和個體化醫療具有重要意義。
在農業和畜牧業中,遺傳變異是育種計劃的基礎。育種者努力保持足夠的遺傳變異,以確保作物和家畜能夠適應未來的環境變化。同時,他們也需要警惕小群體育種可能導致的遺傳漂變,這可能導致有用性狀的丟失或有害突變的積累。
總之,遺傳變異和遺傳漂變是塑造生物群體遺傳結構的兩個基本過程。遺傳變異提供了進化的原材料,而遺傳漂變則引入了隨機性,特別是在小群體中。理解這兩個過程的相互作用對於解釋自然群體的遺傳結構、預測未來的進化趨勢、制定保護策略和指導育種計劃都具有重要意義。隨著基因組學技術的進步,我們對這些過程的認識正在不斷深化,為生物學研究開闢了新的前景。
適應性進化是生物學中一個核心概念,它描述了生物體如何通過自然選擇過程逐漸變得更適合其環境的現象。這個過程涉及遺傳變異、選擇壓力和時間的相互作用,最終導致生物體在形態、生理或行為上的改變,使其能夠更好地生存和繁衍。
適應性進化的基礎是達爾文提出的自然選擇理論。根據這一理論,具有有利特徵的個體更有可能存活並繁衍後代,從而將這些有利特徵傳遞給下一代。隨著時間的推移,這些有利特徵在群體中變得越來越普遍,最終導致整個物種的改變。
遺傳變異是適應性進化的原材料。沒有遺傳變異,自然選擇就無法發揮作用。遺傳變異主要來源於突變、基因重組和基因流動。突變是DNA序列的改變,可能產生新的等位基因。基因重組發生在有性生殖過程中,通過減數分裂時的交叉互換和配子的隨機結合,產生新的基因組合。基因流動則是指不同群體之間的基因交流,通常通過個體遷移實現。
選擇壓力是驅動適應性進化的關鍵因素。這些壓力可能來自非生物因素(如溫度、濕度、光照等)或生物因素(如捕食者、競爭者、寄生物等)。不同的選擇壓力可能導致不同的適應性特徵。例如,在乾旱環境中,植物可能進化出深根系統或耐旱的葉片結構;而在高捕食壓力下,動物可能進化出更快的奔跑速度或更好的偽裝能力。
適應性進化可以分為幾種主要類型。方向性選擇(directional selection)推動群體朝一個極端發展,如長頸鹿頸部的延長。穩定性選擇(stabilizing selection)則傾向於保持群體的中間表型,減少極端變異,如人類出生體重的相對穩定。分裂選擇(disruptive selection)有利於群體中的兩個極端,可能導致物種的分化,如達爾文雀的喙部大小分化。
適應性進化的速度可以根據選擇壓力的強度和遺傳變異的可用性而變化。在某些情況下,適應性進化可以非常快速,這種現象被稱為快速適應性進化(rapid adaptive evolution)。例如,在工業革命期間,英國的樺尺蛾(peppered moth)迅速從淺色變為深色,以適應被煤煙污染的樹皮。這種快速適應性進化在應對人為環境變化(如抗生素耐藥性)或入侵新棲息地時尤為常見。
然而,適應性進化也面臨著一些限制和權衡。首先,遺傳變異的可用性可能有限。如果群體中缺乏適應所需的遺傳變異,那麼即使面臨強烈的選擇壓力,也無法進行適應性進化。其次,一個特徵的改變可能對其他特徵產生負面影響,這種現象被稱為適應性權衡(adaptive trade-off)。例如,增加肌肉質量可能提高力量,但也會增加能量需求。最後,歷史或發育限制可能阻礙某些進化路徑,這就是所謂的系統發生阻力(phylogenetic constraint)。
適應性進化的研究方法包括野外觀察、實驗室實驗和計算機模擬。野外觀察可以提供自然選擇作用的直接證據,如研究達爾文雀喙部大小隨乾旱年份的變化。實驗室實驗允許研究者控制選擇壓力和追蹤多代的進化過程,這在微生物研究中特別有效。計算機模擬則可以探索長時間尺度或複雜情景下的進化動態。
在分子水準上,適應性進化可以通過比較不同物種或群體的DNA序列來研究。正選擇(positive selection)通常會導致功能基因中非同義替換(改變氨基酸)的比率高於同義替換(不改變氨基酸)的比率。這種比較可以通過計算dN/dS比率(非同義替換率與同義替換率的比值)來量化。如果dN/dS大於1,通常被解釋為正選擇的證據。
適應性進化還涉及基因表達的調控。表觀遺傳修飾(epigenetic modification),如DNA甲基化和組蛋白修飾,可以影響基因的表達而不改變DNA序列。這些修飾可以快速響應環境變化,並且在某些情況下可以跨代遺傳,為適應性進化提供了另一個維度。
生態位適應(niche adaptation)是適應性進化的一個重要方面。它描述了物種如何演化以適應特定的生態位,即在生態系統中的特定角色或位置。這種適應可能涉及形態、生理或行為的改變,使物種能夠更有效地利用特定資源或生存在特定環境條件下。例如,仙人掌通過演化出肉質莖和特殊的光合作用方式,適應了乾旱環境。
協同進化(coevolution)是適應性進化的一種特殊形式,它描述了兩個或多個物種之間的相互適應過程。這種現象在捕食者-獵物、寄生物-宿主、植物-授粉者等關係中尤為常見。例如,某些蘭花和特定的傳粉昆蟲之間的形態互補就是協同進化的結果。
適應性進化在應用生物學中也有重要意義。在農業中,了解作物和病原體的適應性進化有助於開發持久的抗性品種和有效的病蟲害管理策略。在醫學領域,研究病原體的適應性進化對於預測和應對新興傳染病至關重要。在保護生物學中,了解物種的適應能力有助於評估它們面對氣候變化和棲息地破壞的脆弱性。
然而,適應性進化並不總是完美的。有時,歷史遺留的特徵可能不再適應當前環境,這種現象被稱為適應不良(maladaptation)。例如,人類對高熱量食物的偏好在食物稀缺的環境中是有利的,但在現代社會中可能導致肥胖問題。此外,局部適應(local adaptation)可能導致群體在面對環境變化時缺乏靈活性。
適應性進化還涉及一個重要的概念:適應度景觀(fitness landscape)。這是一個理論模型,描述了不同基因型或表型與其適應度之間的關係。在這個模型中,適應度被表示為一個多維空間中的"高度",而進化過程則被視為群體在這個景觀上的"攀爬"。適應度景觀的形狀可能影響進化的速度和方向,例如,一個平滑的景觀可能允許漸進的適應,而一個崎嶇的景觀可能導致跳躍式進化或進化停滯。
在研究適應性進化時,科學家們還關注表型可塑性(phenotypic plasticity)的作用。這是指單一基因型能夠產生不同表型的能力,通常作為對環境變化的反應。高度的表型可塑性可能減緩適應性進化的速度,因為它允許生物體在不改變基因型的情況下應對環境變化。然而,表型可塑性本身也可能是適應性進化的結果,特別是在變化多端的環境中。
適應性進化的研究還需要考慮權衡和限制。一個性狀的改善可能以牺牲另一個性狀為代價,這種現象被稱為進化權衡(evolutionary trade-off)。例如,在植物中,增加種子大小可能提高幼苗的存活率,但同時減少種子的數量。此外,發育和物理限制可能限制某些進化路徑的可能性,這就是所謂的進化約束(evolutionary constraint)。
最後,值得注意的是,適應性進化不應被誤解為朝著某個預定目標或"更高級"形式的進化。相反,它是生物體對當前環境條件的持續響應。今天看似完美的適應可能在未來的環境中變得不利。因此,適應性進化是一個動態的、持續的過程,反映了生命與其不斷變化的環境之間的複雜互動。
適應性進化是生物學中的一個核心概念,它描述了生物體如何通過自然選擇的過程逐漸變得更適合其環境的現象。這個過程涉及遺傳變異、選擇壓力和時間的相互作用,最終導致生物體在形態、生理或行為上的改變,使其能夠更好地生存和繁衍。適應性進化是達爾文進化論的核心,也是現代綜合進化論的基石之一。
適應性進化的基礎是群體中存在的遺傳變異。這些變異主要來源於突變、基因重組和基因流動。突變是DNA序列的改變,可能產生新的等位基因。雖然大多數突變是中性或有害的,但偶爾也會出現有益突變,為適應性進化提供原材料。基因重組發生在有性生殖過程中,通過減數分裂時的交叉互換和配子的隨機結合,產生新的基因組合。這種重組可以將有利的等位基因組合在一起,增加適應的可能性。基因流動則是指不同群體之間的基因交流,通常通過個體遷移實現,可以引入新的遺傳變異。
選擇壓力是驅動適應性進化的關鍵因素。這些壓力可能來自非生物因素(如溫度、濕度、光照等)或生物因素(如捕食者、競爭者、寄生物等)。不同的選擇壓力可能導致不同的適應性特徵。例如,在乾旱環境中,植物可能進化出深根系統或耐旱的葉片結構;而在高捕食壓力下,動物可能進化出更快的奔跑速度或更好的偽裝能力。選擇壓力的強度和持續時間會影響適應性進化的速度和程度。
適應性進化可以通過幾種不同的選擇模式進行。方向性選擇(directional selection)推動群體朝一個極端發展,如長頸鹿頸部的延長。在這種情況下,具有較長頸部的個體能夠獲取更多食物,因此有更高的生存和繁殖機會。穩定性選擇(stabilizing selection)則傾向於保持群體的中間表型,減少極端變異。例如,人類出生體重的相對穩定就是穩定性選擇的結果,因為過輕或過重的嬰兒都有較高的死亡風險。分裂選擇(disruptive selection)有利於群體中的兩個極端,可能導致物種的分化。達爾文雀的喙部大小分化就是一個典型的例子,不同大小的喙適應於不同大小的種子。
適應性進化的速度可以根據選擇壓力的強度和遺傳變異的可用性而變化。在某些情況下,適應性進化可以非常快速,這種現象被稱為快速適應性進化(rapid adaptive evolution)。一個經典的例子是英國工業革命期間的樺尺蛾(peppered moth)。隨著空氣污染使樹皮變黑,淺色的樺尺蛾容易被捕食者發現,而深色個體則具有生存優勢。在短短幾十年內,樺尺蛾群體就從以淺色為主變為以深色為主。這種快速適應性進化在應對人為環境變化(如抗生素耐藥性)或入侵新棲息地時尤為常見。
然而,適應性進化也面臨著一些限制和權衡。首先,遺傳變異的可用性可能有限。如果群體中缺乏適應所需的遺傳變異,那麼即使面臨強烈的選擇壓力,也無法進行適應性進化。這就是為什麼保護瀕危物種的遺傳多樣性如此重要。其次,一個特徵的改變可能對其他特徵產生負面影響,這種現象被稱為適應性權衡(adaptive trade-off)。例如,增加肌肉質量可能提高力量,但也會增加能量需求。最後,歷史或發育限制可能阻礙某些進化路徑,這就是所謂的系統發生阻力(phylogenetic constraint)。例如,哺乳動物的脊椎數量相對固定,這限制了頸部長度的進化潛力。
適應性進化的研究方法包括野外觀察、實驗室實驗和計算機模擬。野外觀察可以提供自然選擇作用的直接證據。例如,彼得和羅斯瑪麗·格蘭特(Peter and Rosemary Grant)對加拉帕戈斯群島上達爾文雀的長期研究,記錄了雀鳥喙部大小如何隨乾旱年份的變化而改變。實驗室實驗允許研究者控制選擇壓力和追蹤多代的進化過程,這在微生物研究中特別有效。理查德·倫斯基(Richard Lenski)的大腸桿菌長期進化實驗就是一個著名的例子,展示了細菌如何在新環境中適應和進化。計算機模擬則可以探索長時間尺度或複雜情景下的進化動態,特別是在無法進行實際實驗的情況下。
在分子水準上,適應性進化可以通過比較不同物種或群體的DNA序列來研究。正選擇(positive selection)通常會導致功能基因中非同義替換(改變氨基酸)的比率高於同義替換(不改變氨基酸)的比率。這種比較可以通過計算dN/dS比率(非同義替換率與同義替換率的比值)來量化。如果dN/dS大於1,通常被解釋為正選擇的證據。這種方法已經被廣泛用於識別人類進化中受到正選擇的基因,如與高原適應相關的EPAS1基因。
適應性進化還涉及基因表達的調控。表觀遺傳修飾(epigenetic modification),如DNA甲基化和組蛋白修飾,可以影響基因的表達而不改變DNA序列。這些修飾可以快速響應環境變化,並且在某些情況下可以跨代遺傳,為適應性進化提供了另一個維度。例如,研究發現某些植物在遭受病原體攻擊後,可以通過表觀遺傳修飾來"記住"這種經歷,並將這種抗性傳遞給後代。
生態位適應(niche adaptation)是適應性進化的一個重要方面。它描述了物種如何演化以適應特定的生態位,即在生態系統中的特定角色或位置。這種適應可能涉及形態、生理或行為的改變,使物種能夠更有效地利用特定資源或生存在特定環境條件下。例如,仙人掌通過演化出肉質莖和特殊的光合作用方式(CAM光合作用),適應了乾旱環境。同樣,深海魚類進化出特殊的眼睛結構和發光器官,以適應黑暗的深海環境。
協同進化(coevolution)是適應性進化的一種特殊形式,它描述了兩個或多個物種之間的相互適應過程。這種現象在捕食者-獵物、寄生物-宿主、植物-授粉者等關係中尤為常見。例如,某些蘭花和特定的傳粉昆蟲之間的形態互補就是協同進化的結果。花的形狀和昆蟲的口器結構相互匹配,確保了有效的授粉。另一個著名的例子是豆科植物與根瘤菌之間的共生關係,這種關係通過長期的協同進化形成,使雙方都獲得了生存優勢。
適應性進化在應用生物學中也有重要意義。在農業中,了解作物和病原體的適應性進化有助於開發持久的抗性品種和有效的病蟲害管理策略。例如,通過了解小麥銹病菌的進化模式,科學家們可以預測新的毒性株系的出現,並相應地調整育種策略。在醫學領域,研究病原體的適應性進化對於預測和應對新興傳染病至關重要。HIV病毒的快速進化就是一個挑戰,需要不斷開發新的抗病毒藥物。在保護生物學中,了解物種的適應能力有助於評估它們面對氣候變化和棲息地破壞的脆弱性,從而制定更有效的保護策略。
然而,適應性進化並不總是完美的。有時,歷史遺留的特徵可能不再適應當前環境,這種現象被稱為適應不良(maladaptation)。例如,人類對高熱量食物的偏好在食物稀缺的環境中是有利的,但在現代社會中可能導致肥胖問題。此外,局部適應(local adaptation)可能導致群體在面對環境變化時缺乏靈活性。例如,某些高度特化的物種可能無法應對快速的氣候變化。
適應性進化的研究還需要考慮表型可塑性(phenotypic plasticity)的作用。這是指單一基因型能夠產生不同表型的能力,通常作為對環境變化的反應。高度的表型可塑性可能減緩適應性進化的速度,因為它允許生物體在不改變基因型的情況下應對環境變化。然而,表型可塑性本身也可能是適應性進化的結果,特別是在變化多端的環境中。例如,某些植物可以根據光照條件調整葉片的形態和生理特性,這種可塑性使它們能夠在不同的光環境中生存。
適應性進化的概念也引發了一些哲學和倫理問題。例如,在人類社會中,某些遺傳性疾病的高發可能是過去適應性進化的結果。鐮刀型貧血症就是一個例子,攜帶一個鐮刀型血紅蛋白基因的個體對瘧疾有抵抗力,但兩個基因的個體則會患上嚴重的貧血症。這種情況引發了關於人類干預自然選擇過程的討論。
在結束這個話題時,值得強調的是,適應性進化是一個持續的過程,沒有最終的"完美"狀態。生物體總是在不斷變化的環境中尋求平衡,而今天的適應可能成為明天的負擔。因此,保持遺傳多樣性和生態系統的完整性對於物種的長期生存至關重要。通過深入理解適應性進化的機制和模式,我們不僅能更好地理解生命的歷史和現狀,還能為未來的生物多樣性保護和可持續發展提供科學指導。
系統發生與生命歷史的研究是生物學中的一個重要領域,它旨在揭示不同生物物種之間的進化關係以及生命在地球上的發展歷程。這個領域結合了古生物學、分子生物學、生態學和遺傳學等多個學科的知識,為我們理解生物多樣性的起源和演變提供了重要的理論框架。
系統發生學(Phylogenetics)是研究生物進化關係的科學。它試圖重建生物物種之間的演化歷史,並以系統發生樹(phylogenetic tree)的形式來表示這些關係。系統發生樹是一種分支圖,顯示了不同物種或更高分類單位之間的親緣關係。樹的每個分支代表一個分類群,分支的分叉點代表共同祖先。通過分析系統發生樹,科學家們可以推斷出物種之間的親緣關係,以及特定性狀或基因的進化歷史。
構建系統發生樹的方法主要有兩類:基於形態特徵的方法和基於分子序列的方法。傳統上,科學家們主要依靠對生物體外部和內部形態特徵的比較來推斷親緣關係。這種方法在研究現存生物和化石生物時都很有用。然而,形態特徵有時可能會產生誤導,因為趨同進化(convergent evolution)可能導致不相關的物種發展出相似的特徵。
隨著分子生物學技術的發展,基於DNA或蛋白質序列的系統發生分析變得越來越重要。這種方法假設,序列中的差異反映了物種分歧後經過的時間。通過比較不同物種的同源基因序列,可以推斷出它們的親緣關係。分子鐘(molecular clock)理論進一步假設,在某些基因中,突變以相對恒定的速率積累。這使得科學家們可以估計物種分歧的時間。
然而,分子系統發生學也面臨一些挑戰。例如,基因水準轉移(horizontal gene transfer),特別是在細菌和古菌中常見,可能會干擾系統發生分析。此外,不同基因可能有不同的進化速率,這就需要仔細選擇合適的基因進行分析。因此,現代的系統發生研究通常結合了形態學和分子生物學的方法,以獲得更全面和可靠的結果。
生命歷史(history of life)研究則關注生命在地球上的起源和演化過程。這個領域綜合了來自地質學、古生物學、生物化學和遺傳學等多個學科的證據。根據目前的科學共識,生命可能起源於大約38-41億年前。最早的生命形式可能是簡單的單細胞生物,類似於今天的細菌或古菌。
地球早期的環境與今天大不相同。原始大氣中缺乏氧氣,海洋中充滿了各種無機和有機化合物。在這樣的環境中,簡單的有機分子可能通過化學反應逐漸形成更複雜的分子,最終形成能夠自我複製的系統。這個假說被稱為化學進化(chemical evolution)。然而,從無生命物質到最簡單的生命形式的過渡仍然是科學界熱烈討論的話題。
生命歷史的早期階段主要由原核生物主導。在地球歷史的前30億年左右,生命主要以細菌和古菌的形式存在。這些微生物逐漸改變了地球的環境,最顯著的例子就是光合細菌釋放氧氣,導致大氣中氧氣含量的增加。這個過程被稱為大氧化事件(Great Oxygenation Event),發生在約24億年前。
真核生物的出現是生命歷史上的一個重要里程碑。根據內共生理論(endosymbiosis theory),真核細胞可能起源於原核生物之間的共生關係。例如,線粒體可能起源於被吞噬的原始細菌,而葉綠體可能起源於光合細菌。真核生物的出現為更複雜的多細胞生物的演化鋪平了道路。
多細胞生物的演化是生命歷史中另一個重要的階段。最早的多細胞生物化石可以追溯到大約6億年前的埃迪卡拉生物群(Ediacaran biota)。然而,生物多樣性的大爆發發生在寒武紀早期,約5.4億年前。這個被稱為寒武紀大爆發(Cambrian explosion)的事件見證了幾乎所有現代動物門類的出現。
自寒武紀以來,生命經歷了多次的輻射適應和大滅絕事件。輻射適應(adaptive radiation)是指一個祖先物種快速分化為多個新物種以適應不同生態位的過程。例如,達爾文雀在加拉帕戈斯群島上的輻射適應就是一個經典案例。大滅絕事件(mass extinction events)則導致大量物種在相對短的時間內滅絕。歷史上最嚴重的大滅絕事件發生在二疊紀末期,約2.5億年前,導致超過90%的海洋物種和70%的陸地脊椎動物滅絕。
陸地生物的演化是生命歷史中另一個重要的主題。植物和動物從水生環境向陸地遷移是一個漫長而複雜的過程。最早的陸地植物可能出現在約4.7億年前,而最早的陸地動物可能出現在約4.2億年前。這些早期陸地生物面臨著許多挑戰,如防止脫水、支撐身體、呼吸空氣等。因此,它們演化出了許多適應陸地生活的特徵,如角質層、維管組織、肺部等。
哺乳動物和鳥類的演化是脊椎動物進化史上的重要篇章。哺乳動物起源於約2.25億年前的早期獸形類爬行動物。它們在恐龍統治地球期間經歷了漫長的演化過程,主要以小型夜行動物的形式存在。恐龍滅絕後,哺乳動物經歷了快速的輻射適應,演化出了多種多樣的形態和生活方式。鳥類則被認為是恐龍的後裔,最早的鳥類化石可以追溯到約1.5億年前。
人類的演化是生命歷史研究中最引人入勝的話題之一。人類屬(Homo)的起源可以追溯到約250-300萬年前的非洲。通過對化石記錄和基因組數據的研究,科學家們重建了人類演化的大致過程,包括直立行走、腦容量增大、工具使用等關鍵特徵的出現。現代人類(Homo sapiens)約在20-30萬年前出現在非洲,隨後遷移到世界各地。
系統發生學和生命歷史研究不僅幫助我們理解生物多樣性的起源和演變,還為許多實際應用提供了理論基礎。例如,在醫學領域,系統發生分析被用來追蹤病原體的起源和傳播,指導疫苗的開發。在保護生物學中,了解物種的演化歷史有助於制定更有效的保護策略。在農業中,系統發生學知識被用於作物和家畜的改良。
然而,系統發生與生命歷史研究也面臨著一些挑戰。化石記錄的不完整性使得某些演化事件的精確時間和順序難以確定。分子數據的解釋有時也存在爭議,特別是當不同基因或不同分析方法得出矛盾結果時。此外,隨著新技術和新方法的發展,一些長期被接受的觀點可能需要修正。
儘管如此,系統發生與生命歷史研究仍然是現代生物學中最活躍和最富成果的領域之一。它不僅幫助我們理解生命的過去,還為預測生物對未來環境變化的反應提供了重要依據。隨著研究方法的不斷改進和新證據的不斷湧現,我們對生命演化過程的理解也在不斷深化。
系統發生樹是生物學中用來表示不同物種或生物群體之間進化關係的圖形表示方法。它以樹狀結構呈現,反映了生物之間的親緣關係和演化歷史。系統發生樹的概念最早可以追溯到達爾文的進化論,但隨著現代分子生物學和計算技術的發展,它已經成為了研究生物多樣性和進化的重要工具。
系統發生樹的基本結構包括幾個關鍵元素。樹的頂端稱為根(root),代表所有研究對象的共同祖先。樹的末端稱為葉(leaves)或末端分類單元(terminal taxa),代表現存的物種或研究對象。連接根和葉的線稱為分支(branches),代表進化的路徑。分支的交叉點稱為節點(nodes),代表假設的共同祖先。兩個相鄰節點之間的距離稱為分支長度(branch length),通常用來表示進化時間或遺傳變異的程度。
構建系統發生樹的方法主要分為兩大類:基於距離的方法和基於字符的方法。基於距離的方法首先計算每對分類單元之間的遺傳距離,然後根據這些距離構建樹。最常用的基於距離的方法包括鄰接法(Neighbor-Joining)和UPGMA(Unweighted Pair Group Method with Arithmetic Mean)。這些方法計算速度快,適合處理大量數據,但可能會丟失一些細節信息。
基於字符的方法則直接分析每個字符(如DNA序列中的每個位點)的狀態。最大似然法(Maximum Likelihood)和貝葉斯推斷(Bayesian Inference)是兩種常用的基於字符的方法。這些方法通過評估不同樹拓撲結構的可能性來選擇最佳的樹。雖然計算量較大,但這些方法通常能提供更準確的結果和更詳細的進化信息。
系統發生樹的可靠性評估是一個重要問題。最常用的方法是自展法(bootstrap),它通過反覆從原始數據中有放回地抽樣並重建樹,然後計算每個分支出現的頻率。自展值高的分支被認為是可靠的。另一種方法是後驗概率(posterior probability),常用於貝葉斯分析中,它直接給出了特定分支存在的概率。
解讀系統發生樹需要注意幾個關鍵點。首先,樹的拓撲結構比分支的具體排列更重要。兩個形狀不同但拓撲結構相同的樹實際上表達了相同的進化關係。其次,除非特別說明,分支的長短通常代表進化變化的多少,而不是時間的長短。再者,現存物種(樹的葉)之間的親緣關係應該通過追溯到它們的最近共同祖先來判斷,而不是簡單地看它們在樹上的距離。
系統發生樹在生物學研究中有廣泛的應用。在分類學中,它為物種的分類和命名提供了理論基礎,支持了系統分類學的發展。在進化生物學中,系統發生樹幫助研究者追蹤特定性狀或基因的進化歷史,理解適應性進化和趨同進化等現象。在生態學中,系統發生樹被用來研究群落的形成過程和生物多樣性的分佈模式。
在醫學和公共衛生領域,系統發生樹分析被廣泛用於研究病原體的進化和傳播。例如,在新冠病毒大流行期間,科學家們通過構建病毒基因組的系統發生樹,追蹤了病毒的起源和傳播路徑,為疫情防控提供了重要依據。類似地,系統發生分析也被用於研究抗藥性的演化,幫助開發更有效的抗生素和抗病毒藥物。
系統發生樹還在保護生物學中發揮重要作用。通過分析瀕危物種的系統發生關係,研究者可以評估生物多樣性的丟失程度,並制定更有效的保護策略。例如,系統發生多樣性(phylogenetic diversity)的概念被提出用來量化一個群落或生態系統中的進化歷史多樣性,這為保護決策提供了新的視角。
在分子進化研究中,系統發生樹是理解基因家族演化的重要工具。通過比較不同物種中同源基因的系統發生關係,研究者可以推斷基因複製、功能分化和丟失的過程。這種分析對於理解基因功能的演化和預測未知基因的功能都有重要意義。
系統發生樹的構建和分析也面臨一些挑戰。長分支吸引(long branch attraction)是一個常見問題,它可能導致進化速率快的分支被錯誤地聚在一起。不完全譜系分選(incomplete lineage sorting)是另一個複雜因素,它發生在物種形成的早期階段,可能導致基因樹和物種樹的不一致。水準基因轉移(horizontal gene transfer),特別是在細菌和古菌中普遍存在,也可能干擾系統發生分析。
為了解決這些問題,研究者開發了多種策略。例如,使用多基因或全基因組數據可以減少單個基因帶來的偏差。考慮異質性進化速率的模型可以緩解長分支吸引問題。種系樹方法(species tree methods)則試圖整合多個基因樹的信息來推斷物種樹,以解決不完全譜系分選的問題。
隨著測序技術的進步和大數據分析方法的發展,系統發生樹分析正在向更大規模和更高精度的方向發展。全基因組系統發生學(phylogenomics)使得研究者能夠基於整個基因組的信息來構建進化樹,這大大提高了系統發生推斷的準確性,尤其是對於解決一些長期存在爭議的進化關係問題。
時間校正的系統發生樹(time-calibrated phylogenetic trees),也稱為年代發生樹(chronograms),是另一個重要發展方向。這種樹不僅顯示了物種之間的親緣關係,還提供了各分支分歧時間的估計。通過結合化石記錄和分子鐘方法,研究者可以重建生物進化的時間框架,這對於理解生物多樣性的歷史動態和預測未來的演化趨勢都有重要意義。
系統發生樹的視覺化和註釋也在不斷改進。交互式的系統發生樹可視化工具允許研究者和公眾更直觀地探索複雜的進化關係。將形態、生態或地理信息映射到系統發生樹上,可以幫助研究者發現性狀演化或生物地理學的模式。
在教育領域,系統發生樹已成為教授進化理論的重要工具。它幫助學生直觀地理解物種之間的關係和生物多樣性的形成過程。然而,研究表明許多學生和公眾在解讀系統發生樹時仍然存在困難,這突顯了加強進化教育的重要性。
系統發生樹分析也正在其他學科中找到應用。在語言學中,研究者使用類似的方法來研究語言的演化和關係。在考古學中,系統發生方法被用來分析文化特徵的傳播和演變。這些跨學科的應用展示了系統發生思維的普適性。
儘管系統發生樹分析已經取得了巨大進展,但仍然存在一些未解決的問題和爭議。例如,如何最好地整合形態學和分子數據仍然是一個活躍的研究領域。如何處理網狀進化(reticulate evolution),如雜交和基因渙散(introgression),也是當前研究的熱點。此外,如何在保持計算效率的同時處理越來越大的數據集也是一個持續的挑戰。
分類學是生物學中的一個基礎學科,專門研究生物的命名、描述和分類。這門學科的歷史可以追溯到古希臘時期,但現代分類學的基礎是由18世紀的瑞典博物學家卡爾·林奈奠定的。林奈提出了二名法(binomial nomenclature)系統,即用屬名和種加詞來命名每一個物種,這一系統至今仍在使用。分類學不僅為生物多樣性研究提供了基本框架,也為其他生物學分支如生態學、進化生物學和保護生物學提供了重要支持。
分類學的基本單位是物種(species)。然而,物種的定義一直是生物學中的一個複雜問題。傳統上,生物學家使用形態學物種概念,即根據生物體的外觀和結構來界定物種。隨後,恩斯特·邁爾提出了生物學物種概念,將物種定義為可以相互交配並產生可育後代的群體。近年來,隨著分子生物學技術的發展,系統發生學物種概念和基因型團簇物種概念等新的物種定義也被提出。這些不同的物種概念反映了生物多樣性的複雜性,也為分類學帶來了挑戰。
在林奈的系統中,生物被分為界(kingdom)、門(phylum)、綱(class)、目(order)、科(family)、屬(genus)和種(species)七個主要等級。隨著對生物多樣性認識的深入,分類學家還增加了一些中間等級,如亞界(subkingdom)、超門(superphylum)等。每個分類等級都有其特定的命名規則,這些規則被收錄在國際命名法規(International Code of Nomenclature)中。例如,屬名必須是單詞且首字母大寫,而種加詞通常是形容詞或名詞且小寫。
傳統的分類方法主要依賴於形態學特徵。分類學家通過觀察和比較生物體的外部和內部結構來確定它們之間的相似性和差異。這種方法在植物和動物分類中仍然廣泛使用。然而,形態學方法也有其局限性。例如,它難以區分形態相似但遺傳上不同的隱存種(cryptic species),也難以處理形態變異大的物種。
隨著分子生物學技術的發展,DNA條形碼技術(DNA barcoding)成為了分類學的重要工具。這種方法使用標準化的DNA片段(如動物中的細胞色素氧化酶I基因)來識別和區分物種。DNA條形碼技術特別適用於識別形態學難以區分的物種,如昆蟲幼蟲或微生物。然而,這種方法也存在一些限制,如線粒體基因的母系遺傳可能導致雜交物種的誤判。
整合分類學(integrative taxonomy)是近年來發展起來的一種綜合方法,它結合了形態學、分子生物學、生態學和生物地理學等多種證據來界定物種。這種方法能夠提供更全面和可靠的分類結果,特別是在處理複雜的分類問題時。例如,在研究熱帶雨林中的昆蟲多樣性時,研究者可能會結合形態特徵、DNA序列、生態位偏好和地理分佈等多種信息來確定物種界限。
分類學與系統學(systematics)密切相關但有所不同。分類學主要關注生物的命名和分類,而系統學則更注重研究生物之間的進化關係。現代分類學越來越多地採用系統發生分類法(phylogenetic taxonomy),即根據生物的進化關係來進行分類。這種方法試圖使分類系統反映真實的進化歷史,從而使分類更加自然和有意義。
分類學在生物學研究中扮演著基礎性的角色。在生態學研究中,準確的物種鑒定是理解群落結構和生態系統功能的前提。在保護生物學中,分類學為瀕危物種的識別和保護提供了基礎。在醫學和農業領域,分類學對於識別病原體、害蟲和有益生物至關重要。此外,分類學還為生物多樣性編目和監測提供了必要的工具和方法。
然而,分類學也面臨著一些挑戰。首先是分類學家短缺的問題。隨著老一代分類學家的退休,培養新一代具有專業分類知識的科學家變得越來越重要。其次是未描述物種的巨大數量。儘管科學家已經命名和描述了數百萬種生物,但估計仍有大量物種尚未被發現,特別是在熱帶雨林、深海和土壤微生物群落中。此外,全球氣候變化和棲息地破壞正在加速生物多樣性的喪失,這使得分類學工作變得更加緊迫。
為了應對這些挑戰,分類學正在經歷數字化革命。數字化標本庫和在線物種數據庫使得研究者能夠更容易地獲取和共享分類信息。例如,全球生物多樣性信息設施(Global Biodiversity Information Facility, GBIF)提供了數億條物種分佈記錄,極大地促進了生物多樣性研究。人工智能和機器學習技術也正在被應用於物種識別和分類,這有望加速新物種的發現和描述過程。
分類學還在不斷擴展其研究範圍。例如,微生物組學(microbiomics)的發展為研究難以培養的微生物提供了新的方法。環境DNA(environmental DNA, eDNA)技術允許研究者通過分析環境樣本中的DNA來檢測物種的存在,這對於監測稀有或隱蔽物種特別有用。此外,功能性狀分類法(functional trait taxonomy)試圖將物種的生態功能整合到分類系統中,這對於理解生態系統功能和預測環境變化的影響具有重要意義。
在教育和公眾參與方面,分類學也在尋求創新。公民科學項目,如 iNaturalist 平台,允許業餘愛好者參與物種觀察和鑒定,這不僅有助於收集大量生物多樣性數據,也提高了公眾的自然意識。虛擬和增強現實技術正被用於創建交互式的物種識別工具,使學習分類學變得更加直觀和有趣。
分類學的發展也反映了我們對生命本質理解的深化。早期的分類系統主要基於可觀察的形態特徵,後來納入了解剖學和胚胎學證據,現在則越來越多地依賴於分子和基因組數據。這種演變反映了我們對生物相似性和差異性理解的進步。然而,如何平衡和整合這些不同類型的證據仍然是分類學中的一個重要問題。
在實際應用中,分類學為許多領域提供了重要支持。在農業中,準確的害蟲和病原體鑒定對於作物保護至關重要。在食品安全領域,分類學技術被用於檢測食品摻假和識別非法野生動物製品。在法醫學中,分類學方法可以幫助確定生物證據的來源。在藥物研發中,分類學為尋找新的藥用植物和微生物提供了指導。
分類學還在應對全球變化帶來的挑戰。氣候變化正在改變物種的分佈範圍,有時會導致形態或行為的變化,這為物種界定和分類帶來了新的複雜性。入侵物種的快速擴散也要求分類學家能夠迅速識別和描述這些新的入侵者。此外,隨著極地冰蓋融化和深海探索的推進,分類學家正在發現和描述大量新的極地和深海物種。
在生物多樣性保護中,分類學發揮著關鍵作用。準確的物種鑒定和分類是評估物種瀕危狀態、制定保護策略和監測保護效果的基礎。例如,IUCN紅色名錄(IUCN Red List)依賴於準確的分類信息來評估物種的保護狀況。此外,分類學還幫助識別生物多樣性熱點和特有物種,為保護區的規劃和管理提供科學依據。
同源特徵和類同特徵是生物學中兩個重要的概念,它們在理解生物進化和分類中扮演著關鍵角色。這兩個概念反映了生物特徵的不同進化起源,對於正確解釋生物之間的關係至關重要。
同源特徵(Homologous traits)是指不同生物體中由共同祖先遺傳下來的特徵。這些特徵可能在外觀和功能上有所不同,但它們擁有相同的發育起源和基本結構。例如,人類的手臂、鯨魚的鰭肢和蝙蝠的翅膀都是同源器官。儘管它們的外觀和功能各不相同,但它們都起源於共同祖先的前肢。這種共同的進化起源使得同源特徵成為研究生物親緣關係的重要依據。
同源特徵的識別通常基於幾個關鍵標準。首先是相似的解剖結構和發育模式。例如,所有脊椎動物的前肢都遵循相同的基本骨骼結構:一個上臂骨,兩個前臂骨,和多個腕骨和指骨。其次是相似的胚胎發育過程。比如,所有脊椎動物的前肢都是從體側芽(lateral bud)發育而來。最後,相關基因的表達模式也可以提供同源性的證據。許多控制肢體發育的基因,如HOX基因,在不同脊椎動物中高度保守。
同源特徵可以進一步分為同功同源和異功同源兩種。同功同源(homology)指的是保持原有功能的同源特徵,如人類和黑猩猩的手。異功同源(homoplasy)則是指功能發生改變的同源特徵,如鯨魚的鰭肢和蝙蝠的翅膀。異功同源特徵反映了生物對不同環境的適應,是理解適應性進化的重要線索。
相比之下,類同特徵(Analogous traits)是指不同生物體中外觀或功能相似,但沒有共同進化起源的特徵。這些特徵通常是因為生物面臨相似的環境壓力而獨立進化出來的。類同特徵是趨同進化(convergent evolution)的結果,反映了不同生物對相似環境的適應。
類同特徵的一個經典例子是鳥類和昆蟲的翅膀。儘管它們都用於飛行,但它們的結構和起源完全不同。鳥類的翅膀是由前肢演化而來,而昆蟲的翅膀則是由體壁外突演化而來。另一個例子是魚類和海豚的流線型身體。儘管它們都適應了水生環境,但魚類是真正的水生動物,而海豚則是由陸生哺乳動物演化而來。
識別類同特徵通常需要仔細的比較解剖學和發育生物學研究。雖然表面上看起來相似,但深入研究往往會揭示結構上的顯著差異。例如,章魚和脊椎動物的眼睛表面上非常相似,都能形成清晰的圖像。然而,它們的內部結構和發育過程完全不同,反映了它們是獨立進化的結果。
理解同源和類同特徵對於構建準確的生物系統發生關係至關重要。同源特徵提供了生物之間共同祖先的證據,因此是建立系統發生樹的重要依據。相反,類同特徵可能誤導分類學家,使不相關的物種被錯誤地歸為一類。因此,區分同源和類同特徵是進行系統發生分析的關鍵步驟。
在分子生物學時代,同源和類同的概念也被應用到基因和蛋白質水準。同源基因(homologous genes)是指由共同祖先基因演化而來的基因。它們可以進一步分為直系同源基因(orthologous genes)和旁系同源基因(paralogous genes)。直系同源基因是由物種分化事件產生的,而旁系同源基因則是由基因複製事件產生的。理解這些關係對於推斷基因功能和重建基因家族的進化歷史非常重要。
類似地,類同基因(analogous genes)是指在不同物種中獨立進化出相似功能的基因。這種現象在微生物中特別常見,因為它們面臨的選擇壓力可能導致不相關的基因獲得相似的功能。識別類同基因對於理解生物的適應性進化和功能趨同非常重要。
在進化發育生物學(Evolutionary Developmental Biology,簡稱Evo-Devo)中,同源和類同的概念提供了理解形態進化的重要框架。例如,研究表明許多形態上的創新實際上是由現有發育基因的調控變化引起的,而不是新基因的出現。這解釋了為什麼形態上差異巨大的物種可能共享許多相同的發育基因。
同源特徵還可以幫助我們理解生物的歷史。例如,鯨魚骨骼中存在的退化後肢骨骼是其陸生祖先的證據。類似地,人類尾骨的存在反映了我們與有尾靈長類的共同祖先。這些被稱為痕跡器官(vestigial organs)的結構提供了物種進化歷史的重要線索。
另一方面,類同特徵展示了自然選擇的力量和生物適應性的靈活性。例如,仙人掌和多肉植物雖然屬於不同的科,但都進化出了儲水的肉質莖和葉,這反映了它們對乾旱環境的適應。同樣,不同綱的動物如鯊魚(軟骨魚綱)和海豚(哺乳綱)都進化出了流線型的身體和背鰭,這是對水生環境的適應。
在實際的生物學研究中,區分同源和類同特徵有時可能具有挑戰性。某些特徵可能既有同源成分又有類同成分。例如,鳥類和蝙蝠的翅膀在骨骼結構上是同源的(都起源於四足動物的前肢),但它們用於飛行的適應性卻是獨立進化的結果。這種情況被稱為平行進化(parallel evolution),它介於純粹的同源和純粹的類同之間。
分子生物學技術的發展為識別同源和類同特徵提供了新的工具。通過比較不同物種的基因序列和表達模式,研究者可以更準確地判斷特徵的進化起源。例如,即使兩個特徵在形態上看起來完全不同,如果它們由相同的基因控制,那麼它們很可能是同源的。相反,如果形態相似的特徵由不同的基因控制,那麼它們可能是類同的。
然而,即使在分子水準上,區分同源和類同有時也不是直截了當的。例如,趨同進化可能導致不相關的序列獲得相似的功能,這被稱為分子趨同(molecular convergence)。同樣,基因的水準轉移(horizontal gene transfer),特別是在微生物中,可能使不相關的物種獲得相似的基因,這可能被誤解為同源關係。
理解同源和類同特徵對於許多應用領域都有重要意義。在藥物開發中,了解不同物種間蛋白質的同源關係可以幫助研究者預測藥物的作用和副作用。在農業中,識別作物中的同源基因可以指導育種策略,幫助開發具有理想特性的新品種。在生態學研究中,理解類同特徵可以幫助解釋不同物種如何共存和競爭。
同源和類同特徵的概念也深刻影響了我們對生物進化的理解。它們展示了進化既有其連續性(通過同源特徵體現),又有其創新性(通過類同特徵體現)。這種理解不僅豐富了我們對生物多樣性起源的認識,也為预測生物对未来环境变化的响应提供了理论基础。
支序分類學是一種生物分類方法,它基於共同祖先的原則來確定生物之間的進化關係。這種方法最初由德國昆蟲學家威利·亨尼希(Willi Hennig)在20世紀50年代提出,並在隨後的幾十年中逐漸成為系統分類學的主導方法。支序分類學的核心思想是通過識別共有衍生特徵(synapomorphy)來確定生物群體之間的親緣關係,並將這些關係以支序圖(cladogram)的形式呈現出來。
支序分類學的基本單位是單系群(monophyletic group)或支序群(clade),它包括一個共同祖先及其所有後代。這與傳統分類學中常用的並系群(paraphyletic group)和多系群(polyphyletic group)形成對比。並系群包括一個共同祖先但不包括其所有後代,而多系群則包括沒有最近共同祖先的成員。支序分類學堅持只有單系群才是有效的分類單位,這一原則極大地改變了生物分類的方式。
在支序分類學分析中,研究者首先需要確定外群(outgroup)和內群(ingroup)。外群是與研究對象相對較遠的親緣群體,用於確定特徵的祖先狀態。內群則是研究的目標群體。然後,研究者會收集一系列可能反映進化關係的特徵,這些特徵可以是形態學的、生理學的、行為學的,也可以是分子水準的,如DNA或蛋白質序列。
支序分類學使用特徵狀態的轉變來推斷進化關係。原始狀態(plesiomorphic state)是祖先型的特徵狀態,而衍生狀態(apomorphic state)是由原始狀態演變而來的新狀態。共有衍生特徵是支序分類學中最重要的概念之一,它指的是一個群體共同擁有的衍生特徵,這種特徵表明該群體有共同的祖先。例如,所有哺乳動物都有乳腺,這是一個共有衍生特徵,表明所有哺乳動物都來自一個共同的祖先。
支序分類學分析的結果通常以支序圖的形式呈現。支序圖是一種樹狀圖,每個分支代表一個分類群,分支的分叉點代表共同祖先。支序圖的每個節點都代表一個共有衍生特徵的出現。重要的是,支序圖只顯示相對的分支順序,而不表示進化時間或分支間的遺傳距離。
支序分類學的一個核心原則是節約原則(parsimony principle)。這個原則假設進化過程中特徵狀態的改變次數最少的解釋是最可能的。因此,在構建支序圖時,研究者會尋找需要最少進化步驟的樹狀結構。這種方法被稱為最大節約法(maximum parsimony method)。
然而,隨著分子數據的廣泛應用,其他方法如最大似然法(maximum likelihood method)和貝葉斯推斷(Bayesian inference)也被引入支序分類學分析中。這些方法考慮了更複雜的進化模型,可以處理大量的分子數據,並提供統計支持的估計。
支序分類學的應用極大地改變了我們對生物分類和進化的理解。例如,基於支序分類學的分析,鳥類被認為是恐龍的直接後代,實際上是恐龍的一個分支。這導致了分類系統的重大調整,許多傳統的分類群被重新定義或廢棄。例如,"爬行動物"這個傳統分類群現在被認為是一個並系群,因為它不包括鳥類,而鳥類實際上是爬行動物的一個分支。
支序分類學也為研究生物特徵的進化提供了框架。通過在支序圖上映射特徵,研究者可以推斷特徵的演化歷史,識別平行進化和趨同進化的情況。這種方法被稱為特徵狀態重建(character state reconstruction),它幫助我們理解特徵是如何隨時間演變的,以及哪些因素可能驅動了這種演變。
在分子生物學領域,支序分類學的原理被用於研究基因家族的演化。通過構建基因樹,研究者可以識別基因複製、丟失和功能分化的事件。這對於理解基因功能的演化和預測未知基因的功能都有重要意義。例如,通過比較不同物種的同源基因,研究者可以推斷基因功能的保守性和多樣性。
支序分類學也為生物地理學研究提供了工具。通過比較生物的系統發生關係和地理分佈,研究者可以推斷物種的起源地和擴散路徑。這種方法被稱為支序地理學(cladistic biogeography),它幫助我們理解地質事件如何影響了生物的進化和分佈。
然而,支序分類學也面臨一些挑戰。首先是同源性評估的問題。確定不同生物的特徵是否真正同源有時可能很困難,特別是當面對高度衍生或退化的結構時。其次,趨同進化和平行進化可能導致不相關的生物具有相似的特徵,這可能誤導分類分析。
另一個挑戰是如何處理網狀進化(reticulate evolution)的情況。支序分類學假設進化是分歧的過程,但在某些生物群中,尤其是植物和微生物中,雜交和基因水準轉移是常見的現象。這些過程可能導致基因譜系和物種譜系的不一致,使得構建單一的支序圖變得困難。
為了解決這些問題,研究者開發了一些新的方法和工具。例如,網絡方法(network methods)被用來表示複雜的進化關係,包括可能的雜交事件。基因組水準的分析(genomic-level analysis)可以提供更全面的進化信息,幫助解決單個基因可能帶來的偏差。多基因連鎖分析(multi-gene concatenation analysis)和種系樹方法(species tree methods)則試圖整合來自多個基因的信息來推斷物種樹。
支序分類學的原理和方法也被應用到生物學以外的領域。在語言學中,類似的方法被用來研究語言的演化和關係。在考古學中,支序分類學的思想被用來分析文化特徵的傳播和演變。這些跨學科的應用展示了支序思維的普適性。
在教育領域,支序分類學為教授進化理論提供了有力的工具。通過構建和解釋支序圖,學生可以直觀地理解物種之間的關係和生物多樣性的形成過程。然而,研究表明許多學生和公眾在理解支序圖時仍然存在困難,這突顯了加強進化教育的重要性。
支序分類學還在生物信息學領域發揮重要作用。隨著測序技術的進步和大數據分析方法的發展,研究者可以基於全基因組數據進行大規模的系統發生分析。這種全基因組支序分類學(genome-wide cladistics)為解決一些長期存在爭議的進化關係問題提供了新的機會。
在保護生物學中,支序分類學的方法被用來評估生物多樣性的程度和價值。系統發生多樣性(phylogenetic diversity)是一個基於支序分類學原理的指標,它考慮了物種之間的進化關係。這種方法可以幫助確定具有獨特進化歷史的物種或群體,為保護決策提供科學依據。
支序分類學還促進了對生物適應性和特徵演化的理解。通過在支序圖上映射生態和形態特徵,研究者可以推斷特徵的演化速率和方向,識別可能的適應性輻射事件。這種方法被稱為比較方法(comparative method),它為研究適應性進化提供了強大的工具。
總的支序分類學不僅革新了生物分類的方法,也為理解生物進化提供了全新的視角。它強調了基於共同祖先的分類原則,促進了系統分類學與進化生物學的緊密結合。儘管仍然面臨一些挑戰,但隨著新技術和方法的不斷發展,支序分類學將繼續在生物學研究中發揮重要作用。
系統發生樹是生物學中用來表示物種或其他分類單位之間進化關係的圖形模型。這種樹狀結構不僅反映了生物之間的親緣關係,也展示了生命的歷史和多樣性。然而,隨著研究的深入和新技術的應用,科學家們對系統發生樹的理解和詮釋也在不斷evolve發展。這些不同的觀點豐富了我們對生命進化過程的認識,同時也帶來了新的挑戰和機遇。
傳統的系統發生樹觀點將生命的進化視為一個分歧的過程,像樹的分支一樣,物種從共同祖先分化出來,形成新的類群。這種觀點強調了物種之間的階層關係和共同祖先的重要性。在這個模型中,每個分支點代表一個假設的共同祖先,而樹的頂端則代表現存的物種或分類群。這種樹狀模型直觀且易於理解,因此在生物學教育和研究中被廣泛採用。
然而,隨著分子生物學的發展,科學家們發現生命的進化過程比原先想像的要複雜得多。水準基因轉移(Horizontal Gene Transfer, HGT)的發現對傳統的樹狀模型提出了挑戰。HGT是指遺傳物質在不同物種之間直接轉移的現象,這在細菌和古菌中特別普遍。這意味著基因的傳遞不僅僅是從親代到子代的垂直過程,還包括物種之間的橫向交流。這種現象使得某些生物群體的進化歷史更像是一個網絡,而不是簡單的樹狀結構。
為了適應這種複雜性,科學家們提出了網狀進化(Reticulate Evolution)的概念。網狀進化模型試圖捕捉基因流動的複雜性,包括物種之間的雜交、基因滲入和HGT等現象。這種模型不再將進化描繪為一個簡單的分歧過程,而是承認了物種之間可能存在的複雜基因交流。網狀進化模型特別適用於研究植物和微生物的演化,因為這些生物群體中基因交換的現象更為普遍。
另一個影響系統發生樹觀點的因素是不完全譜系分選(Incomplete Lineage Sorting, ILS)。ILS發生在種群分化的早期階段,當祖先種群中的多態性在新物種形成過程中未能完全分選。這可能導致基因樹與物種樹不一致的情況。ILS的存在意味著即使在沒有基因流動的情況下,不同基因也可能顯示不同的進化歷史。這種現象在快速輻射進化的類群中特別常見,如鳥類和靈長類。
為了處理ILS帶來的挑戰,科學家們開發了種系樹方法(Species Tree Methods)。這些方法試圖從多個基因樹中推斷出一個統一的物種樹。種系樹方法承認單個基因可能有不同的歷史,但仍然尋求重建整個基因組的總體進化歷史。這種方法的應用大大提高了系統發生推斷的準確性,特別是在處理近期快速分化的類群時。
隨著全基因組測序技術的發展,系統發生基因組學(Phylogenomics)成為了研究進化關係的新方法。這種方法利用整個基因組的信息來構建系統發生樹,而不僅僅依賴於少數幾個基因。系統發生基因組學的優勢在於它可以提供更全面的進化圖景,減少單個基因可能帶來的偏差。然而,處理如此大量的數據也帶來了計算和解釋上的挑戰。
時間校正的系統發生樹(Time-Calibrated Phylogenetic Trees),也稱為年代發生樹(Chronograms),為進化研究增添了時間維度。這種樹不僅顯示了物種之間的親緣關係,還估計了各分支分歧的具體時間。時間校正通常結合化石記錄和分子鐘方法來實現。這種方法使得研究者能夠將生物的進化與地質和氣候事件聯繫起來,從而更好地理解驅動物種形成和滅絕的因素。
微生物的系統發生研究帶來了獨特的挑戰和見解。由於微生物的形態相對簡單,傳統的形態學分類方法在這個領域顯得不足。分子方法,特別是16S rRNA基因的分析,革新了微生物的系統發生研究。然而,微生物中普遍存在的HGT現象使得構建一個單一的、全面的微生物系統發生樹變得困難。一些科學家甚至質疑是否存在一個統一的微生物生命之樹。
為了應對這些挑戰,一些研究者提出了核心基因組(Core Genome)的概念。這種方法專注於在特定生物群中高度保守且不太可能經歷HGT的基因。通過分析這些核心基因,研究者試圖重建微生物的基本進化框架。同時,研究者也開發了各種計算方法來識別和量化HGT的影響,以更全面地理解微生物的進化動態。
系統發生樹的另一個重要發展是多基因族系統發生(Multi-Gene Family Phylogenies)的研究。基因複製是生物進化的一個重要機制,它可以產生具有新功能的基因。多基因族系統發生研究不僅關注物種之間的關係,還關注基因家族內部的演化動態。這種方法可以揭示基因功能的演化、基因丟失事件,以及基因家族擴張與物種適應之間的關係。
系統發生樹的視覺化和註釋也在不斷改進。交互式的系統發生樹可視化工具允許研究者和公眾更直觀地探索複雜的進化關係。將形態、生態或地理信息映射到系統發生樹上,可以幫助研究者發現性狀進化或生物地理學的模式。這種整合的方法為理解生物多樣性的起源和維持機制提供了新的視角。
在保護生物學中,系統發生樹的應用帶來了新的思路。系統發生多樣性(Phylogenetic Diversity)的概念被提出用來量化一個群落或生態系統中的進化歷史多樣性。這種方法認為,保護具有獨特進化歷史的物種或群體對於維持生態系統的長期穩定性和適應潛力更為重要。這為保護決策提供了一個新的標準,鼓勵我們不僅關注物種的數量,還要考慮它們所代表的進化歷史。
系統發生樹還在醫學研究中發揮重要作用。在流行病學中,研究者使用系統發生方法來追蹤病原體的起源和傳播路徑。例如,在新冠病毒大流行期間,病毒基因組的系統發生分析幫助科學家理解了病毒的演化速率和傳播動態。在癌症研究中,系統發生方法被用來研究腫瘤內的細胞異質性和癌症進展的動態過程。
然而,系統發生樹的構建和解釋仍然面臨著一些挑戰。長分支吸引(Long Branch Attraction)是一個常見的問題,它可能導致進化速率快的分支被錯誤地聚在一起。為了解決這個問題,研究者開發了各種統計方法和模型選擇技術。另一個挑戰是如何處理缺失數據。在大規模的系統發生基因組學研究中,並非所有物種都有完整的基因組數據。如何在保持分析準確性的同時最大化納入的物種數量,是一個持續的研究課題。
隨著人工智能和機器學習技術的發展,這些方法也開始應用於系統發生分析。機器學習算法可以幫助識別複雜的進化模式,預測基因功能,甚至輔助系統發生樹的構建。這些新技術的應用有望提高系統發生分析的效率和準確性,特別是在處理大規模數據集時。
總的對系統發生樹的不同觀點反映了我們對生命進化過程理解的不斷深化。從簡單的分歧模型到複雜的網絡結構,從單基因分析到全基因組方法,系統發生研究正在揭示生命歷史的豐富性和複雜性。這些進展不僅挑戰了我們對生命樹的傳統看法,也為理解和保護生物多樣性提供了新的工具和視角。隨著研究方法的不斷創新和跨學科合作的加強,我們對生命進化的認識將會更加全面和深入。