 永續社 www.epa.url.tw 林雨莊
永續社 www.epa.url.tw 林雨莊植物的繁殖方式多樣化,依據植物的類型和生長環境,植物可通過有性繁殖和無性繁殖兩種方式進行繁殖。這些繁殖方式不僅確保了植物物種的延續,也促進了植物的適應與演化。
無性繁殖是一種不涉及配子結合的繁殖方式,這種繁殖方式通常發生在植物的體細胞中。無性繁殖的主要特徵是後代與親代遺傳信息相同,這樣的繁殖方式效率較高,能夠迅速擴大植物群體。常見的無性繁殖方式包括分裂、出芽、匍匐莖和地下莖。分裂是指植物體直接分裂成兩個或更多個體,每個個體都能夠獨立生存。出芽則是在親代植物的某一部分上形成芽體,隨著芽體的生長,它最終會脫離母體成為獨立的植物。匍匐莖和地下莖也是植物進行無性繁殖的常見方式,例如草莓的匍匐莖可以沿地表向外生長,形成新的植株;馬鈴薯的地下莖(塊莖)則能在適合的環境中發芽成長。這些無性繁殖方式通常發生在環境相對穩定的情況下,能夠快速增殖。
有性繁殖涉及兩個不同性別的配子的結合,這一過程能夠產生基因多樣性的後代。有性繁殖對植物適應環境變化、進行遺傳重組具有重要意義。植物的有性繁殖最典型的形式發生在種子植物中,這包括了裸子植物和被子植物。裸子植物如松樹和銀杏,其種子裸露在毬果中,並通過風力進行傳播。被子植物則是目前地球上最繁盛的植物類群,其繁殖結構——花,極大地促進了植物繁殖的成功率。
被子植物的花是其有性繁殖的關鍵部位,花朵中的雄蕊和雌蕊分別負責產生花粉和卵細胞。雄蕊的花藥會釋放花粉,這些花粉通過風、昆蟲、鳥類或其他動物傳播到另一朵花的雌蕊上,完成授粉過程。授粉後,花粉中的精子細胞會通過花粉管進入雌蕊的胚珠中,與卵細胞結合,形成受精卵。受精卵發育成胚,周圍的胚珠組織則會轉變為種子,種子的外層則由果實包裹,以保護種子並幫助其傳播。
植物的授粉方式多樣,依賴於不同的傳粉媒介。有些植物依靠風力傳播花粉,這些植物的花通常缺乏鮮豔的色彩或香味,花粉顆粒較輕,便於隨風飄散。風媒花如松樹、榖類植物,這些植物的花粉量大,以確保在隨機的風力傳播中有足夠的花粉能夠到達雌蕊。而其他植物則依賴於動物傳粉,如蜜蜂、蝴蝶、鳥類和蝙蝠等,這些植物會發展出鮮豔的花朵和強烈的香氣,以吸引傳粉者。動物在尋找花蜜時,無意中將花粉攜帶到另一朵花上,完成授粉。這種與動物的互利共生關係極大地提升了授粉的效率和精確度,尤其是在植物物種多樣化和生態環境復雜的區域。
受精後,種子的發育過程也非常重要。種子內部的胚胎由根、胚軸和子葉構成,外層則被堅韌的種皮保護。種子能夠在不利的環境中休眠,直到環境條件適宜時再萌發。種子萌發過程中,水分和適當的溫度是關鍵條件,水分使種皮軟化,胚內的酶系統開始作用,儲存在胚乳或子葉中的養分被轉化為可供胚胎生長的能量。
果實是被子植物另一個重要的繁殖結構,它不僅保護種子,還有助於種子的擴散。不同類型的果實有不同的傳播方式,這些方式包括風力、動物和水等。例如,蒲公英的果實輕盈,帶有冠毛,能夠隨風遠距離傳播;椰子的果實能夠漂浮在水面上,適應海洋或河流環境;而一些果實則會通過吸引動物食用,藉此傳播種子,動物在吃掉果肉後,種子會通過糞便排出,並在其他地方萌發。
植物的繁殖策略也因其生活環境的不同而各具特點。某些植物能夠同時進行有性和無性繁殖,這讓它們在不同的環境壓力下能夠靈活選擇最適合的繁殖方式。這些策略的演化不僅促進了植物在地球上各種環境中的擴散,也促進了植物的多樣性發展。
總之,植物繁殖系統展示了高度的複雜性和多樣性,從無性繁殖的快速擴展到有性繁殖的基因多樣性,每種繁殖方式都有其適應環境的優勢。無論是通過風媒、動物傳粉,還是果實的種子傳播,植物的繁殖機制在確保物種存續和適應環境變化方面發揮了關鍵作用,並成為植物界持續繁榮和多樣化的根本驅動力。
植物生殖是生物學中一個極為重要的領域,對於理解生命的延續和生態系統的平衡至關重要。以下將從多個角度詳細闡述植物生殖的重要性:
首先,植物生殖是維持物種存續的關鍵過程。通過有性和無性生殖,植物得以產生後代,確保基因的傳遞和物種的延續。這一過程不僅保證了植物自身的繁衍,也為依賴植物生存的其他生物提供了持續的食物和棲息地來源。在生態系統中,植物作為初級生產者,其生殖能力直接影響著整個食物鏈的穩定性和生物多樣性。
其次,植物生殖在農業生產中扮演著核心角色。人類的糧食安全和經濟發展在很大程度上依賴於對植物生殖過程的理解和控制。通過選擇性育種和雜交技術,農業科學家能夠培育出產量更高、抗病性更強、營養價值更豐富的作物品種。這不僅提高了農業生產效率,也為解決全球糧食短缺問題提供了可能性。此外,園藝和觀賞植物產業也高度依賴於植物生殖技術,以滿足人們對美化環境和改善生活質量的需求。
植物生殖還在生物技術和醫學領域中具有重要應用。通過對植物生殖機制的深入研究,科學家們開發出了諸如基因工程和克隆等先進技術。這些技術不僅用於改良作物品種,還被應用於生產藥用植物和開發新型藥物。例如,利用植物生殖系統生產疫苗和治療性蛋白質,為人類健康做出了重要貢獻。
從進化生物學的角度來看,植物生殖系統的研究為我們理解生命的起源和演化提供了寶貴的洞察。植物在地球上的演化歷程中,發展出了多樣化的生殖策略,包括從最簡單的孢子繁殖到複雜的種子植物生殖系統。這些不同的生殖方式反映了植物如何適應各種環境條件,也揭示了生物進化的一般規律。
在生態保護方面,了解植物生殖對於保護瀕危物種和維護生態平衡具有重要意義。許多植物物種因棲息地破壞、氣候變化等因素面臨滅絕威脅。通過研究這些植物的生殖特性,科學家們可以制定更有效的保護策略,如人工授粉、種子庫建設等,以維護生物多樣性。
植物生殖還與全球氣候變化密切相關。植物的生殖週期受到溫度、光照等環境因素的影響。氣候變化可能導致植物開花時間的改變,進而影響整個生態系統的平衡。研究植物生殖對氣候變化的響應,有助於預測和應對未來可能出現的生態問題。
在教育領域,植物生殖是生物學課程的重要組成部分。通過學習植物的生殖過程,學生不僅能夠理解生命的基本原理,還能培養科學思維和探究精神。這對於培養下一代科學家和提高公眾的科學素養都具有重要意義。
植物生殖在文化和藝術方面也佔有一席之地。許多文學作品、繪畫和音樂都以植物的生長和繁衍為主題,反映了人類對生命循環的思考和對自然美的欣賞。這種文化層面的影響進一步凸顯了植物生殖在人類社會中的重要地位。
最後,植物生殖研究為解決全球性問題提供了新的思路。例如,在應對糧食安全、能源短缺和環境污染等挑戰時,植物生殖技術可能會發揮關鍵作用。通過培育耐旱、高產的作物品種,或開發能夠高效吸收二氧化碳的植物,我們可以為這些問題找到創新的解決方案。
總之,植物生殖是一個涉及面廣、影響深遠的研究領域。它不僅關乎植物自身的存續,還與人類社會的發展、生態環境的保護以及科技創新的推進密切相關。深入理解和研究植物生殖,將為我們應對當前和未來的挑戰提供重要的科學基礎和實踐指導。
植物的生殖發育與結構是植物學中一個重要而複雜的領域,涉及植物生命週期中的關鍵過程和構造。這一領域的研究不僅有助於我們理解植物如何繁衍後代,還為農業生產、生態保護和生物技術的發展提供了重要的理論基礎。
植物的生殖發育過程可以分為有性生殖和無性生殖兩大類。有性生殖涉及配子的形成、受精和新個體的發育,而無性生殖則通過體細胞或特化結構直接產生新個體。這兩種生殖方式在植物界中都廣泛存在,反映了植物在長期進化過程中適應不同環境的策略。
在有性生殖中,植物首先需要發育出特化的生殖器官。對於被子植物而言,這主要指花的形成。花是被子植物特有的生殖器官,其結構複雜,包括花萼、花瓣、雄蕊和雌蕊。花的發育過程受到嚴格的基因調控,涉及多個基因家族的協同作用。例如,ABC模型解釋了花器官身份決定的基本機制,其中A類基因控制萼片和花瓣的發育,B類基因參與花瓣和雄蕊的形成,C類基因則負責雄蕊和心皮的發育。
雄蕊是植物的雄性生殖器官,主要由花絲和花藥組成。花藥內部包含花粉囊,在這裡進行減數分裂和有絲分裂,最終形成花粉粒。花粉粒是雄配子體,其結構包括外壁、內壁和細胞質。外壁通常具有特殊的紋飾,這不僅有助於花粉的傳播,還在物種識別中起重要作用。花粉粒內部含有營養細胞和生殖細胞,後者在花粉管生長過程中進一步分裂形成兩個精子細胞。
雌蕊是植物的雌性生殖器官,由子房、花柱和柱頭組成。子房內部包含胚珠,這是未來種子的前體。胚珠的發育是一個複雜的過程,涉及多次細胞分裂和分化。在胚珠內部,大孢子母細胞經過減數分裂形成四個半數體細胞,其中通常只有一個存活並發育成雌配子體(胚囊)。成熟的胚囊通常包含七個細胞八個核,其中最重要的是位於珠孔端的卵細胞和中央細胞。
授粉是植物有性生殖的關鍵步驟,指的是花粉從雄蕊轉移到雌蕊柱頭的過程。授粉方式多種多樣,包括風媒、蟲媒、鳥媒和水媒等。不同的授粉方式對應著植物的不同適應性特徵。例如,風媒植物通常具有小而輕的花粉,而昆蟲傳粉的植物則可能有鮮艷的花瓣和特殊的氣味來吸引傳粉者。
授粉後,花粉在柱頭上萌發形成花粉管。花粉管的生長是一個高度精確的過程,受到多種信號分子的調控。花粉管沿著花柱生長,最終到達胚珠,將兩個精子細胞釋放到胚囊中。這一過程中,花粉管的定向生長(趨化性)確保了精子能夠準確到達目標位置。
雙受精是被子植物特有的受精方式。在這個過程中,一個精子與卵細胞結合形成合子,另一個精子與中央細胞結合形成三倍體的胚乳核。合子通過一系列有絲分裂和分化最終發育成胚胎,而胚乳核則發育成為滋養胚胎生長的胚乳組織。
種子的發育是植物生殖週期的重要階段。種子通常包含胚胎、胚乳(或子葉)和種皮。胚胎是未來新植株的雛形,包含了根、莖和葉的原始結構。胚乳或子葉為胚胎的早期生長提供營養。種皮則起保護作用,並可能具有特殊結構以助於種子的傳播。
果實是被子植物特有的結構,由子房壁發育而來。果實的主要功能是保護種子並協助種子傳播。根據子房壁的發育方式,果實可分為真果(如蘋果)和假果(如草莓)。果實的形態和結構多種多樣,反映了植物在種子傳播策略上的多樣性適應。
植物的無性生殖方式也十分多樣。常見的無性生殖方式包括分株繁殖、塊莖繁殖、球莖繁殖、鱗莖繁殖等。這些方式使得植物能夠在不形成種子的情況下快速繁衍。無性生殖產生的後代與親本在遺傳上完全相同,這在某些情況下可能是有利的,例如保持優良品種的特性。
植物生殖發育過程中的激素調控是一個重要研究領域。多種植物激素參與調節生殖器官的發育和功能。例如,生長素和細胞分裂素影響花芽的形成和發育,赤黴素則參與花粉發育和果實生長。乙烯在果實成熟過程中起關鍵作用,而脫落酸則與種子休眠有關。
環境因素對植物生殖發育有重要影響。光週期、溫度、水分和營養狀況等都可能影響植物的開花時間和結實情況。例如,許多植物需要經過春化作用(低溫處理)才能開花,這是植物適應季節變化的一種機制。
在農業生產中,對植物生殖發育的理解和控制至關重要。通過調控開花時間、提高授粉效率、改善果實品質等手段,可以顯著提高作物產量和品質。此外,植物生殖生物學的研究成果也廣泛應用於園藝和觀賞植物培育中。
植物生殖發育研究還為生物技術提供了重要工具。例如,花藥培養和胚胎救援技術可用於克服遠緣雜交不親和的問題,而體細胞胚胎發生技術則為植物快速繁殖和遺傳轉化提供了新途徑。
在生態學研究中,植物的生殖策略是理解植物群落動態和進化的重要切入點。不同的生殖方式反映了植物對特定生態位的適應,也影響著植物的分佈範圍和種群結構。
總之,植物生殖發育與結構是一個涵蓋範圍廣泛、內容豐富的研究領域。它不僅關乎植物學的基礎理論,還與農業生產、生態保護、生物技術等多個應用領域密切相關。深入理解植物的生殖過程和結構,對於解決糧食安全、保護生物多樣性、應對氣候變化等全球性挑戰具有重要意義。隨著分子生物學、基因組學等技術的發展,我們對植物生殖發育的認識正在不斷深化,這將為農業和生態領域帶來新的機遇和挑戰。
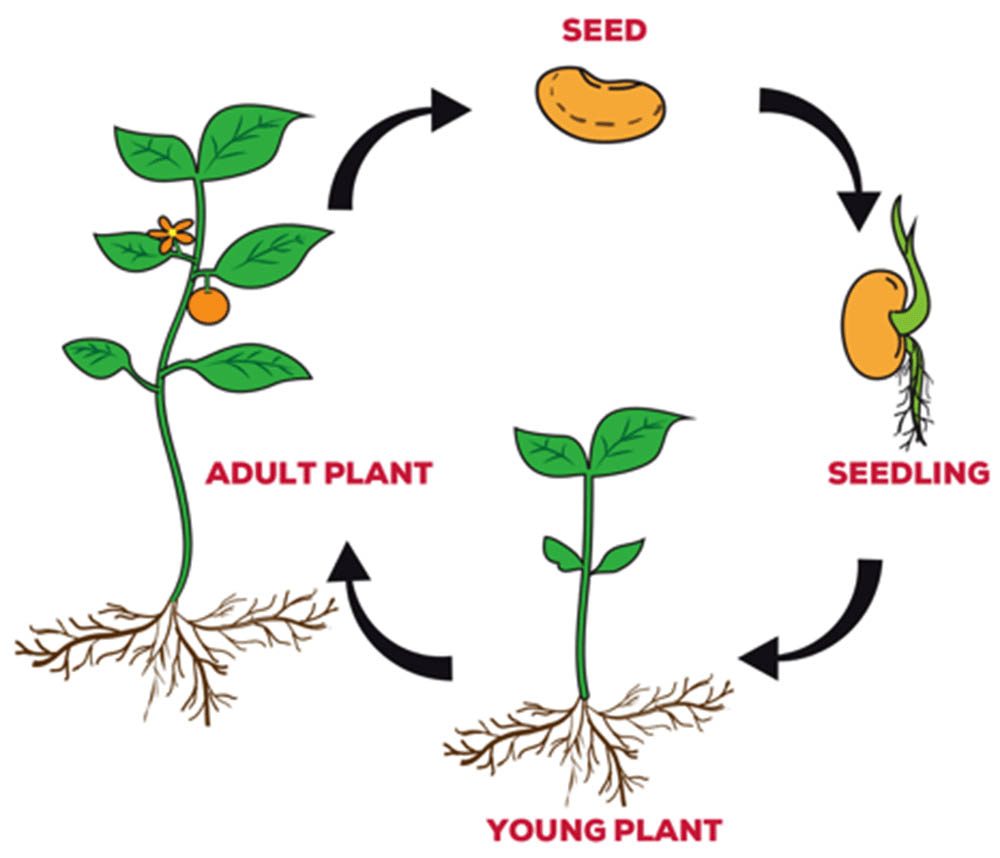
植物的生命週期是一個複雜而精妙的過程,涵蓋了從種子萌發到成熟植株產生新一代種子的全過程。這個週期可以分為幾個主要階段,每個階段都有其特定的生理過程和形態特徵。理解植物生命週期的各個階段對於植物學研究、農業生產和生態保護都具有重要意義。
種子萌發是植物生命週期的起點。種子中的胚胎在適宜的條件下開始生長,這個過程被稱為萌發。萌發的條件通常包括適當的溫度、充足的水分和氧氣。種子首先吸水,激活體內的酶系統。這些酶開始分解儲存的營養物質,為胚胎的生長提供能量和原料。隨後,胚根突破種皮,向下生長形成幼根。胚芽則向上生長,最終突破土壤表面。這個階段的植物完全依賴種子中儲存的養分生存。
幼苗期是植物生命週期的第二個階段。在這個階段,植物開始進行光合作用,逐漸轉為自養生活。幼苗通常有一對子葉(雙子葉植物)或一片子葉(單子葉植物)。這些子葉最初可能仍在為幼苗提供營養,但隨著真葉的出現,光合作用成為植物獲取能量的主要方式。幼苗期的植物非常脆弱,容易受到環境stress的影響,如乾旱、病蟲害等。
營養生長期是植物生命週期中最長的階段。在這個階段,植物主要進行莖、葉和根的生長,不斷增加生物量。莖的生長使植物增高,有助於競爭光照;葉的擴展增加了光合面積;根系的發達則有利於吸收水分和礦物質。植物在這個階段積累的養分為後續的生殖生長做準備。營養生長的速度和持續時間受到多種因素的影響,包括日照長度、溫度、水分和養分供應等。
生殖生長期標誌著植物進入了繁衍後代的階段。對於開花植物這個階段始於花芽的形成。花芽的形成通常受到環境因素的誘導,如日照長度的變化(光週期效應)或溫度的變化(春化作用)。一些植物需要經過一定時期的低溫才能開花,這被稱為春化作用。花芽形成後,植物進入開花期。花的結構複雜,包括花萼、花瓣、雄蕊和雌蕊。雄蕊產生花粉,雌蕊包含胚珠,這些都是植物有性生殖的重要組成部分。
授粉是生殖生長期的關鍵環節。花粉需要從雄蕊轉移到雌蕊的柱頭上,這個過程可以通過風、昆蟲、鳥類等媒介完成。不同的植物種類演化出了不同的授粉策略。例如,風媒植物通常產生大量輕盈的花粉,而昆蟲傳粉的植物則可能有鮮艷的花朵和特殊的氣味來吸引傳粉者。授粉後,花粉在柱頭上萌發,形成花粉管。花粉管沿著花柱生長,最終到達胚珠,將精子細胞釋放到胚囊中。
受精是植物有性生殖的核心過程。被子植物的一個顯著特徵是雙受精。在這個過程中,一個精子與卵細胞結合形成合子,另一個精子與中央細胞結合形成三倍體的胚乳核。合子通過一系列細胞分裂最終發育成胚胎,而胚乳核則發育成為滋養胚胎生長的胚乳組織。這個過程確保了遺傳物質的重組,增加了後代的遺傳多樣性。
種子和果實的發育是植物生命週期的重要階段。受精後,胚珠開始發育成種子。種子通常包含胚胎、儲存組織(如胚乳或子葉)和保護性的種皮。同時,子房壁開始發育成果實。果實的主要功能是保護種子並協助種子傳播。根據植物種類的不同,果實可能是肉質的(如蘋果)或乾燥的(如豆莢)。果實的形態和結構多種多樣,反映了植物在種子傳播策略上的多樣性適應。
種子成熟和散布是植物生命週期的最後階段。成熟的種子通常進入休眠狀態,這使得種子能夠度過不利的環境條件。休眠可能是由種皮的不透水性、胚胎的生理抑制或特定環境條件的缺失等因素造成的。種子的散布方式多種多樣,包括風力傳播、動物傳播、水力傳播等。一些植物的種子具有特殊的結構,如翅膀或鉤刺,以適應特定的傳播方式。
值得注意的是,並非所有植物都完全遵循這個生命週期。一些植物可能會跳過某些階段或以不同的方式完成這些階段。例如,一些多年生植物可能會多次經歷營養生長和生殖生長的循環。此外,許多植物還具有無性繁殖的能力,如通過分株、塊莖或葉芽繁殖等方式產生新的個體。
在植物的生命週期中,世代交替是一個重要的概念。所有植物都經歷配子體和孢子體兩個世代的交替。在蕨類植物中,我們可以清楚地看到這兩個世代:孢子體是我們常見的蕨類植物形態,而配子體則是一個獨立的、通常很小的構造,稱為原葉體。在種子植物中,孢子體世代占主導地位,而配子體世代則大大簡化,雄配子體縮小為花粉粒,雌配子體則縮小為胚囊。
環境因素對植物生命週期的各個階段都有重要影響。例如,溫度和水分條件影響種子的萌發;光照強度和養分供應影響營養生長的速度和程度;日照長度的變化可能誘導開花。了解這些環境因素對植物生命週期的影響,對於農業生產和生態保護都具有重要意義。
在農業生產中,對植物生命週期各階段的控制是提高產量和品質的關鍵。例如,通過調控播種時間來確保作物在最佳條件下萌發和生長;通過施肥和灌溉優化營養生長;通過控制日照時間或使用植物激素來調節開花時間。在果樹栽培中,了解植物的生命週期有助於合理安排修剪、授粉和果實管理等操作。
在生態學研究中,植物的生命週期特徵是理解植物群落動態和演化的重要依據。不同的生活史策略(如年生、二年生或多年生)反映了植物對特定生態位的適應。例如,在資源貧乏或干擾頻繁的環境中,短生命週期的植物可能更具優勢。
隨著分子生物學和遺傳學的發展,我們對控制植物生命週期各階段的基因網絡有了更深入的了解。例如,開花時間的調控涉及多個基因家族的相互作用,包括光週期通路、春化通路和自主開花通路等。這些知識為作物改良提供了新的工具和思路。
總之,植物生命週期的各個階段構成了一個完整而複雜的系統,反映了植物在長期進化過程中對環境的適應。深入理解這些階段及其調控機制,不僅有助於我們更好地認識植物生命的奧秘,也為農業生產和生態保護提供了重要的理論基礎和實踐指導。
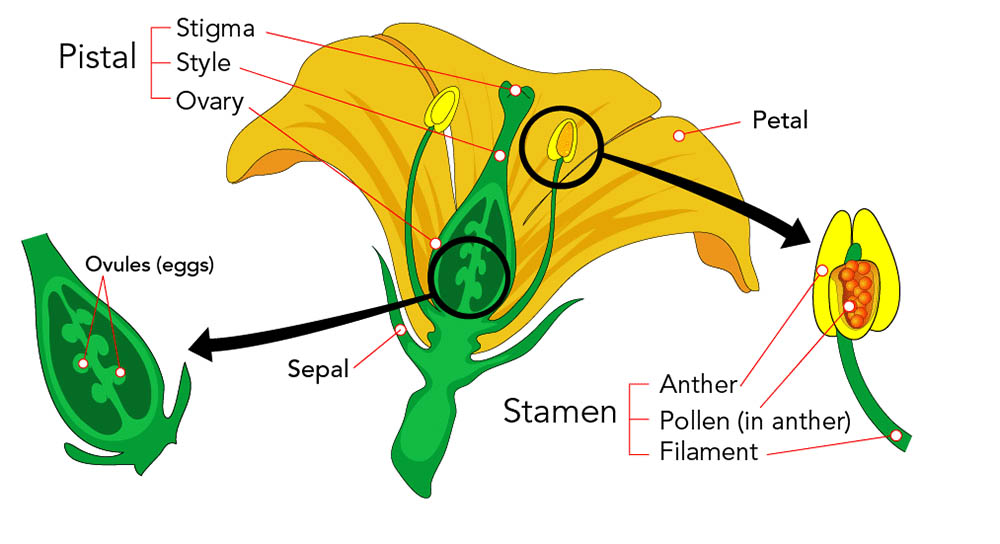
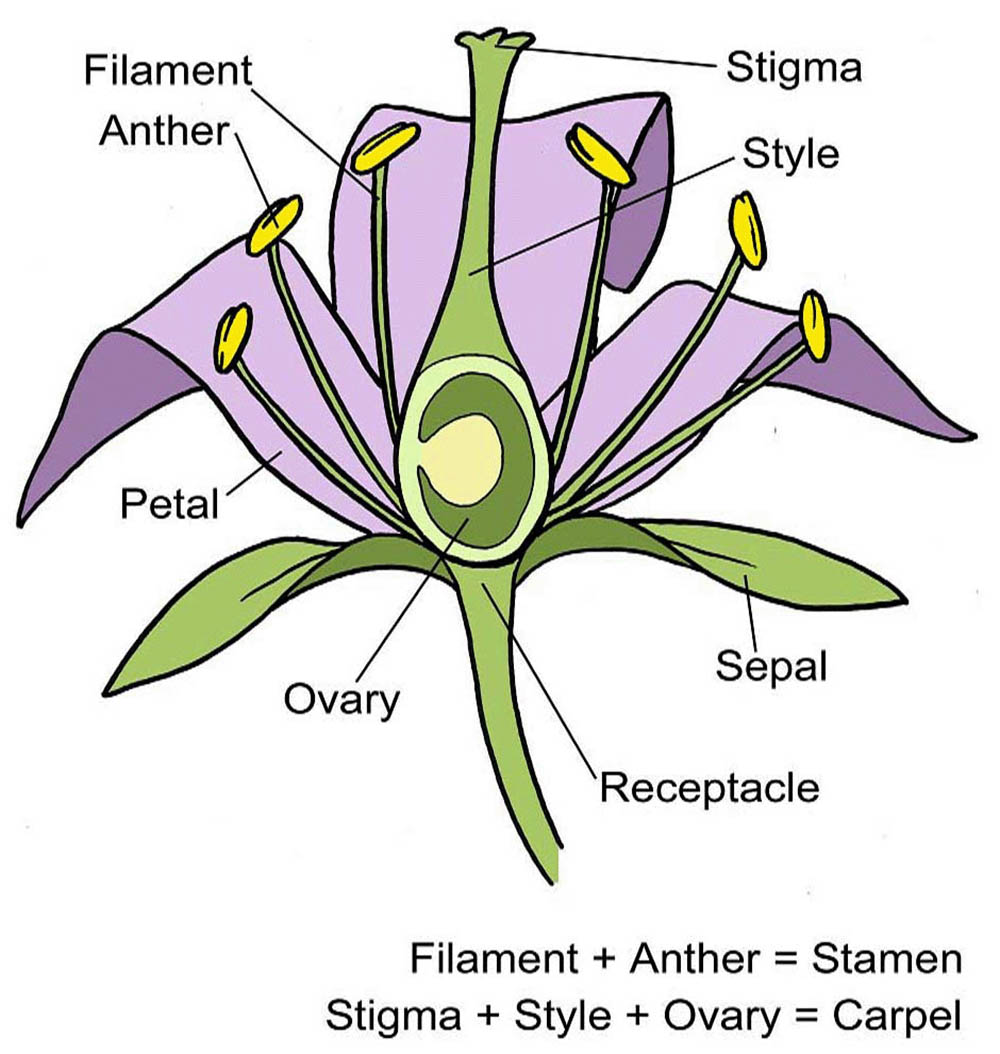
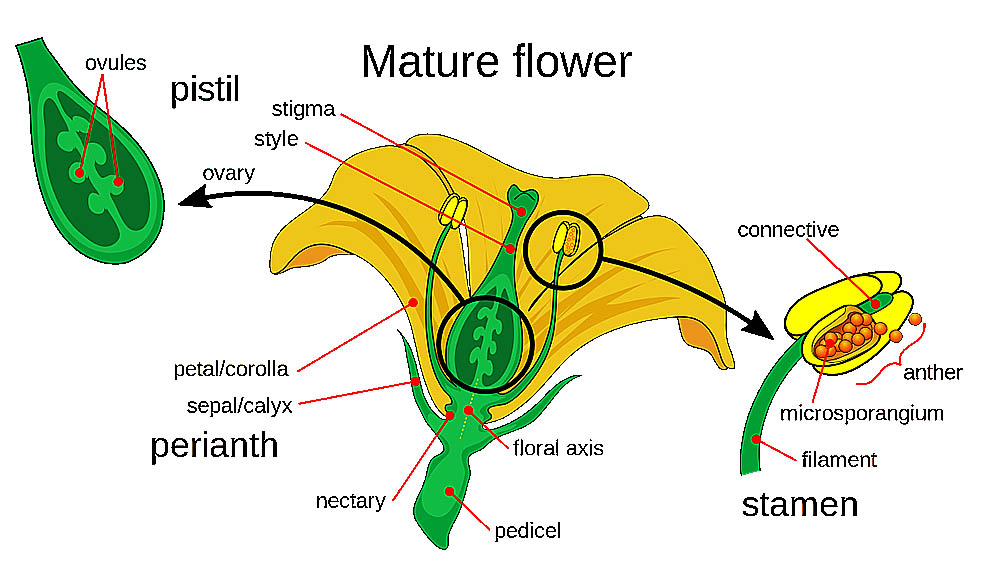
花是被子植物的生殖器官,其結構複雜而精妙,在植物的生命週期中扮演著關鍵角色。花的基本結構可以分為四個主要部分:花萼、花冠、雄蕊和雌蕊。這些部分共同構成了完整的花,每個部分都有其特定的功能和形態特徵。
花萼是花的最外層結構,通常呈綠色,由多片萼片組成。萼片的主要功能是在花蕾階段保護內部的花器官。在某些植物中,花萼可能會變得鮮艷或增大,參與吸引傳粉者的功能。例如,一些熱帶植物的花萼呈現明亮的紅色或黃色,成為視覺上的主要吸引物。花萼的形態多樣,可以是分離的(離萼),也可以部分或完全連合(合萼)。
花冠是花中最引人注目的部分,通常由鮮艷的花瓣組成。花瓣的主要功能是吸引傳粉者,如昆蟲或鳥類。花瓣的顏色、形狀和香氣都是為了適應特定的傳粉者而進化形成的。例如,蝴蝶傳粉的花朵通常有鮮艷的顏色和平台狀的著陸區,而蛾類傳粉的花朵則可能呈現白色並在夜間散發香氣。花瓣的排列方式可以是輻射對稱(如玫瑰)或者兩側對稱(如蘭花)。某些植物的花瓣可能會特化形成複雜的結構,如蘭花的唇瓣。
雄蕊是花的雄性生殖器官,負責產生花粉。一朵花通常有多個雄蕊,每個雄蕊由花絲和花藥兩部分組成。花絲是細長的支持結構,將花藥舉起以便於花粉的傳播。花藥是雄蕊的頂端部分,內含花粉囊,這是花粉形成的場所。花粉囊內進行減數分裂和有絲分裂,最終形成花粉粒。成熟的花粉粒通常具有特殊的外壁結構,這不僅有助於花粉的傳播,還在物種識別中起重要作用。
雌蕊是花的雌性生殖器官,位於花的中心。一朵花可能有一個或多個雌蕊。典型的雌蕊由三個部分組成:柱頭、花柱和子房。柱頭位於雌蕊的頂端,是接收花粉的部位。它通常有粘性或具有特殊的表面結構,以有效地捕獲和保持花粉。花柱連接柱頭和子房,為花粉管的生長提供通道。子房位於雌蕊的基部,內含一個或多個胚珠。胚珠是未來種子的前體,包含了雌配子體(胚囊)。
花的這四個基本部分在不同植物中可能會有顯著的變化。例如,某些植物可能缺少花萼或花冠,而另一些植物可能有額外的結構,如副花冠。花的各個部分的數量和排列方式也可能因植物種類而異,這些特徵常常用於植物分類。
花的對稱性是另一個重要的結構特徵。輻射對稱的花(如蘋果花)從任何角度剖開都能得到相同的兩半,而兩側對稱的花(如豌豆花)只能沿一個平面剖開得到相同的兩半。花的對稱性與其傳粉方式密切相關,例如,兩側對稱的花通常與特定的昆蟲傳粉者相適應。
花的大小也有很大的變化。世界上最大的單花是(拉弗萊西亞)(Rafflesia),其直徑可達一米,而最小的花則來自(瓊麻科)(Wolffia),直徑僅約0.3毫米。花的大小通常與其傳粉方式和生長環境相適應。
某些植物具有特化的花結構,以適應特定的生態位或傳粉方式。例如,(虎耳草)(Saxifraga)的花具有蜜腺盤,專門用於分泌花蜜吸引傳粉昆蟲。而(蘭花科)(Orchidaceae)植物則進化出了極其複雜的花結構,有些甚至模仿特定昆蟲的外形和氣味,以達到精確的傳粉效果。
花的發育過程受到嚴格的基因調控。ABC模型是解釋花器官發育的經典理論,它描述了三類基因(A、B、C類)如何相互作用決定花的四輪同心圓結構。根據這個模型,A類基因單獨表達產生萼片,A類和B類共同表達產生花瓣,B類和C類共同表達產生雄蕊,而C類單獨表達則產生心皮(雌蕊的組成部分)。後來的研究又發現了D類和E類基因,進一步完善了這個模型。
花的結構與其功能密切相關。例如,花粉的產生和傳播、花蜜的分泌、傳粉者的吸引等功能都依賴於特定的花結構。了解花的結構對於理解植物的生殖策略至關重要。例如,自花授粉和異花授粉的植物在花的結構上往往有顯著差異。自花授粉的植物可能有較小的花,雄蕊和雌蕊靠得很近,而異花授粉的植物則可能有較大、較鮮艷的花,雄蕊和雌蕊的位置有利於傳粉者的訪問。
在農業和園藝中,對花的結構的理解有重要的實際應用。例如,在果樹栽培中,了解花的結構有助於判斷開花品質、預測結果率,並指導人工授粉等操作。在觀賞植物的育種中,對花的結構的操縱(如將單瓣花育成重瓣花)是創造新品種的重要手段。
某些植物的花可能會特化形成特殊的聚集體,如(菊科)(Asteraceae)植物的頭狀花序。在這種情況下,多個小花聚集在一起,形成看起來像單朵大花的結構。這種花序結構增加了對傳粉者的吸引力,同時也提高了授粉效率。
花的結構還可能受到環境因素的影響。例如,在資源匱乏的環境中,植物可能會產生較小的花或減少花的數量。氣候變化也可能影響花的結構和開花時間,這可能導致植物與其傳粉者之間的時間錯配,對生態系統產生深遠影響。
在進化生物學研究中,花的結構提供了重要的線索。通過比較不同物種的花結構,科學家可以推斷植物之間的進化關係,了解植物如何適應不同的生態位。例如,風媒傳粉的植物往往有簡化的花結構,缺少鮮艷的花瓣,而具有伸長的雄蕊和羽毛狀的柱頭,這些都是對風媒傳粉的適應。
某些植物可能具有不完全花,即缺少一個或多個典型的花部件。例如,一些植物可能只有雄蕊或只有雌蕊,這種情況被稱為單性花。如果雄花和雌花出現在同一株植物上,稱為雌雄同株;如果出現在不同的植物個體上,則稱為雌雄異株。這些變異反映了植物在繁殖策略上的多樣性。
花的結構研究在植物系統學中扮演著重要角色。花的特徵,如花瓣數量、雄蕊排列方式、子房位置等,常常被用作分類依據。例如,(百合目)(Liliales)的植物通常具有六枚花被片、六枚雄蕊和上位子房,而(蘭科)(Orchidaceae)植物則以其獨特的唇瓣和合蕊柱結構而聞名。
被子植物的有性生殖是一個複雜而精妙的過程,涉及多個階段和多種結構。這個過程不僅確保了遺傳物質的重組和傳遞,也為植物適應不同環境提供了基礎。被子植物的有性生殖過程主要包括花的發育、雄配子體和雌配子體的形成、授粉、受精以及種子和果實的發育。
花的發育是被子植物有性生殖的起點。花是被子植物特有的生殖器官,其發育受到複雜的基因網絡調控。花的發育始於花序分生組織中花原基的形成。隨後,花原基分化形成花的各個部分:萼片、花瓣、雄蕊和雌蕊。這個過程受到所謂的ABC模型基因的調控,其中A類基因控制萼片和花瓣的發育,B類基因參與花瓣和雄蕊的形成,C類基因則負責雄蕊和心皮的發育。後來的研究又發現了D類和E類基因,進一步完善了這個模型。
雄配子體(花粉粒)的形成發生在花藥中。花藥內的小孢子母細胞經過減數分裂形成四分體,每個小孢子隨後發育成花粉粒。成熟的花粉粒通常包含兩個細胞:營養細胞和生殖細胞。營養細胞負責形成花粉管,而生殖細胞在花粉管生長過程中進一步分裂形成兩個精子細胞。花粉粒的外壁通常具有特殊的紋飾,這不僅有助於花粉的傳播,還在物種識別中起重要作用。
雌配子體(胚囊)的形成發生在胚珠中。胚珠內的大孢子母細胞經過減數分裂形成四個大孢子,通常只有一個存活並發育成胚囊。成熟的胚囊通常包含七個細胞八個核,這種結構被稱為(北極型胚囊)(Polygonum type)。其中最重要的是位於珠孔端的卵細胞和中央細胞。卵細胞是未來胚胎的前體,而中央細胞則參與形成胚乳。
授粉是被子植物有性生殖的關鍵步驟,指的是花粉從雄蕊轉移到雌蕊柱頭的過程。授粉方式多種多樣,包括風媒、昆蟲媒介、鳥媒和水媒等。不同的授粉方式對應著植物的不同適應性特徵。例如,風媒植物通常具有小而輕的花粉,而昆蟲傳粉的植物則可能有鮮艷的花瓣和特殊的氣味來吸引傳粉者。某些植物還進化出了複雜的機制來確保異花授粉,如雌雄異熟(雄蕊和雌蕊在不同時間成熟)或自交不親和性。
授粉後,花粉在柱頭上萌發形成花粉管。花粉管的生長是一個高度精確的過程,受到多種信號分子的調控。花粉管沿著花柱生長,最終到達胚珠,將兩個精子細胞釋放到胚囊中。這一過程中,花粉管的定向生長(趨化性)確保了精子能夠準確到達目標位置。花粉管的生長速度非常快,在某些物種中可達每秒幾微米。
雙受精是被子植物特有的受精方式,也是其最顯著的特徵之一。在這個過程中,兩個精子細胞分別與卵細胞和中央細胞結合。一個精子與卵細胞融合形成合子,這是未來胚胎的起源。另一個精子與中央細胞融合形成三倍體的胚乳核,這將發育成為滋養胚胎生長的胚乳組織。雙受精的發現極大地推動了植物學的發展,也為理解被子植物的進化提供了重要線索。
受精後,合子通過一系列有絲分裂和分化最終發育成胚胎。胚胎發育的過程可以分為原胚期、球形胚期、心形胚期、魚雷形胚期等階段。在這個過程中,胚胎逐漸形成了根、莖、葉的原始結構。同時,胚乳也在快速發育,為胚胎提供營養。在某些植物中,胚乳在種子成熟時被完全吸收,營養物質轉移到子葉中(如豆類);而在其他植物中,成熟種子仍保留大量胚乳(如穀類)。
種子的發育是被子植物有性生殖的重要階段。種子通常包含胚胎、胚乳(或子葉)和種皮。種皮由珠被發育而來,起保護作用。某些植物的種子還具有特殊的結構,如翅或冠毛,以助於傳播。種子發育的後期,胚胎進入休眠狀態,代謝活動降到最低水準。這種休眠機制使得種子能夠度過不利的環境條件,等待合適的時機萌發。
果實是被子植物特有的結構,由子房壁發育而來。果實的主要功能是保護種子並協助種子傳播。根據子房壁的發育方式,果實可分為真果(如蘋果)和假果(如草莓)。果實的形態和結構多種多樣,反映了植物在種子傳播策略上的多樣性適應。例如,肉質果實吸引動物食用,從而達到種子傳播的目的;而乾燥的蒴果則可能依靠風力傳播種子。
被子植物的有性生殖過程受到多種內在和外在因素的影響。植物激素在這個過程中起著重要的調控作用。例如,生長素和細胞分裂素影響花芽的形成和發育,赤黴素則參與花粉發育和果實生長。乙烯在果實成熟過程中起關鍵作用,而脫落酸則與種子休眠有關。
環境因素對被子植物的有性生殖也有重要影響。光週期、溫度、水分和營養狀況等都可能影響植物的開花時間和結實情況。例如,許多植物需要經過春化作用(低溫處理)才能開花,這是植物適應季節變化的一種機制。全球氣候變化可能會影響植物的開花時間,進而影響整個生態系統的平衡。
被子植物的有性生殖還表現出許多有趣的適應性特徵。例如,某些植物進化出了自交不親和性機制,防止自花授粉,以增加遺傳多樣性。另一些植物則發展出了特殊的傳粉綜合徵,如(蘭科)(Orchidaceae)植物模仿特定昆蟲的外形和氣味,以實現精確的傳粉。這些適應性特徵反映了植物與環境之間的共同進化。
在農業生產中,對被子植物有性生殖的理解和控制至關重要。通過調控開花時間、提高授粉效率、改善果實品質等手段,可以顯著提高作物產量和品質。例如,在溫室番茄生產中,常使用蜜蜂或振動裝置輔助授粉,以提高坐果率。在果樹栽培中,了解不同品種的自交親和性對於合理搭配授粉樹至關重要。
被子植物的有性生殖研究為生物技術提供了重要工具。例如,通過人工控制授粉,可以實現遠緣雜交,創造新的品種。體外受精技術則為克服生殖隔離提供了可能。此外,對有性生殖過程中關鍵基因的研究,為作物的遺傳改良提供了新的靶點。
被子植物的有性生殖是一個複雜而精妙的過程,涉及多個階段和多種結構。這個過程不僅確保了遺傳物質的重組和傳遞,也為植物適應不同環境提供了基礎。深入理解這一過程,對於基礎植物學研究、農業生產和生態保護都具有重要意義。
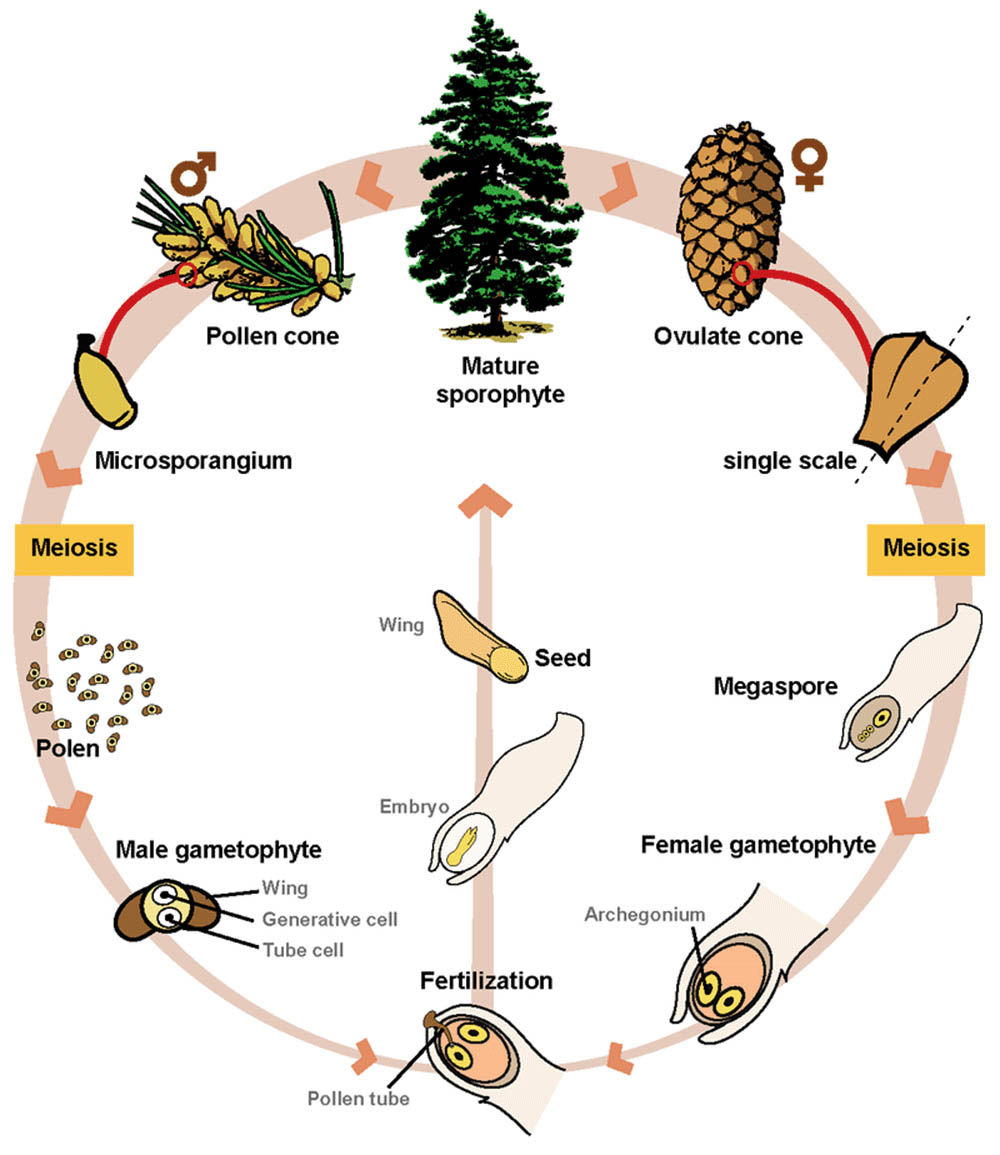
裸子植物的有性生殖過程是植物界中一個獨特而重要的現象。作為種子植物中較為原始的一支,裸子植物的生殖方式既保留了某些古老的特徵,又展現出對陸地生活的適應。裸子植物的有性生殖過程包括配子體的形成、授粉、受精以及種子的發育等多個階段。
裸子植物的最顯著特徵是其"裸露"的種子,這也是其名稱的由來。與被子植物不同,裸子植物的胚珠暴露在空氣中,沒有被子房包裹。這種結構使得裸子植物的授粉和受精過程具有獨特性。
裸子植物的生殖器官主要是球果。雄球果和雌球果可能出現在同一株植物上(雌雄同株),也可能分別位於不同的植物個體上(雌雄異株)。雄球果通常較小,由多個鱗片狀的小孢子葉組成,每個小孢子葉下方有多個花粉囊。雌球果則較大,由多個大孢子葉組成,每個大孢子葉上通常有兩個胚珠。
雄配子體(花粉粒)的形成是裸子植物有性生殖的重要一環。在雄球果的花粉囊中,小孢子母細胞經過減數分裂形成四個單倍體的小孢子。每個小孢子隨後發育成一個花粉粒。裸子植物的花粉粒結構較為特殊,通常包含多個細胞。以松樹為例,成熟的花粉粒包含四個細胞:兩個前孢夫細胞、一個管細胞和一個生殖細胞。其中,前孢夫細胞形成特殊的氣囊,有助於風媒傳播。
雌配子體的發育發生在胚珠內部。胚珠中的大孢子母細胞經過減數分裂形成四個大孢子,通常只有一個存活並發育成雌配子體。裸子植物的雌配子體比被子植物的大得多,通常包含數千個細胞。在雌配子體的頂部形成數個頸卵器,每個頸卵器包含一個卵細胞。這種結構被認為是裸子植物較為原始的特徵之一,與蕨類植物的頸卵器有一定相似性。
授粉是裸子植物有性生殖的關鍵步驟。裸子植物主要依賴風媒傳粉,這與其簡單的花粉結構和暴露的胚珠相適應。風媒傳粉雖然效率較低,但使得裸子植物能夠在大範圍內進行基因交流。當花粉落在胚珠的珠孔上時,授粉完成。
在授粉後,花粉粒開始萌發並形成花粉管。花粉管的生長過程在裸子植物中可能持續很長時間,有些種類甚至需要一年以上。這是裸子植物與被子植物的另一個重要區別。花粉管生長的過程中,生殖細胞分裂形成兩個精子細胞。
受精過程發生在花粉管到達胚珠後。一個精子細胞與卵細胞結合形成合子,這是未來胚胎的起源。與被子植物不同,裸子植物沒有雙受精現象,另一個精子細胞通常會退化。受精後,合子開始分裂發育成胚胎,同時胚乳組織也開始發育。裸子植物的胚乳是單倍體的,這是它們與被子植物的又一個重要區別。
種子的發育是裸子植物有性生殖的最後階段。種子由胚胎、胚乳和種皮組成。胚胎發育過程中形成了根、莖、葉的原始結構。胚乳為胚胎提供營養,在某些裸子植物中,胚乳可能會被完全消耗。種皮由珠被發育而來,起保護作用。某些裸子植物的種子還具有特殊的結構,如松樹種子的翅,有助於風媒傳播。
裸子植物的生殖週期通常較長。以松樹為例,從球果形成到種子成熟可能需要兩到三年的時間。這種長週期的生殖策略使得裸子植物能夠更好地適應環境變化,但也增加了繁殖的風險。
裸子植物的有性生殖過程展現出許多有趣的適應性特徵。例如,某些裸子植物的雌球果分泌黏液來捕獲花粉,這可能是對風媒傳粉低效率的一種補償。另一些裸子植物,如銀杏,其精子具有鞭毛,可以游動到卵細胞處,這被認為是一種古老的特徵。
在裸子植物中,(蘇鐵類)(Cycads)展現出一些獨特的生殖特徵。蘇鐵是現存最古老的種子植物之一,其生殖方式保留了許多原始特徵。蘇鐵的精子具有鞭毛,能夠游動到卵細胞處完成受精。這種特徵在其他現存裸子植物中已經消失,反映了蘇鐵在進化上的保守性。
(松柏類)(Conifers)是裸子植物中最大的一類,包括松樹、杉樹、柏樹等。松柏類的生殖過程展現出對風媒傳粉的高度適應。例如,松樹的花粉具有氣囊,可以在空氣中漂浮很長距離。雌球果通常位於樹冠頂部,有利於接收花粉。某些松樹species的球果需要森林火災的熱量才能開裂釋放種子,這是對特定生態環境的適應。
(銀杏)(Ginkgo)是現存最古老的裸子植物之一,被稱為"活化石"。銀杏的生殖過程保留了許多原始特徵。例如,銀杏的精子具有鞭毛,可以游動到卵細胞處。銀杏的受精過程發生在脫落的種子中,這是一種獨特的適應。銀杏是雌雄異株的,這可能有助於增加遺傳多樣性。
(麻黃類)(Ephedra)是裸子植物中較為特殊的一類,其生殖結構顯示出向被子植物過渡的趨勢。麻黃的雌球果結構簡單,通常只有一個胚珠。其種子被特化的苞片包裹,這在某種程度上類似於被子植物的果實。
裸子植物的有性生殖過程受到多種環境因素的影響。溫度、光照和水分條件都可能影響球果的發育、花粉的釋放和傳播以及種子的成熟。例如,某些松樹species需要經過一段時間的低溫才能誘導球果形成,這是對季節變化的適應。
在演化的角度來看,裸子植物的有性生殖方式代表了植物從孢子繁殖到種子繁殖的重要過渡。種子的出現使得植物能夠更好地適應陸地環境,為被子植物的繁盛奠定了基礎。同時,裸子植物保留的一些原始特徵,如游動的精子、大型的雌配子體等,為我們理解植物的演化歷程提供了重要線索。
裸子植物的有性生殖研究對於林業生產具有重要意義。了解不同樹種的開花結實規律,可以指導種子採集和育苗工作。例如,通過控制環境條件,可以誘導某些經濟樹種提前開花結果,加速育種過程。在森林恢復和生態建設中,了解裸子植物的生殖特性有助於制定更有效的造林策略。
在生態學研究中,裸子植物的有性生殖過程是理解森林生態系統動態的重要切入點。例如,球果和種子的產量變化可能影響依賴這些資源的動物種群,進而影響整個生態系統的平衡。氣候變化可能改變裸子植物的開花結實週期,這對於預測和應對全球變化的生態影響具有重要意義。
裸子植物的有性生殖還為生物技術研究提供了獨特的材料。例如,裸子植物的大型雌配子體為研究植物胚胎發育提供了便利的實驗系統。某些裸子植物種類能夠產生單倍體植株,這為遺傳學研究和育種工作提供了有價值的材料。
在古植物學研究中,化石記錄為我們理解裸子植物有性生殖的演化提供了重要證據。例如,泥盆紀末期出現的種子蕨化石顯示了最早期種子植物的生殖結構,這些化石記錄幫助我們重建了種子植物的演化歷程。
人工控制裸子植物的有性生殖過程在園林綠化和觀賞植物培育中有重要應用。通過人工授粉、嫁接等技術,可以培育出新的園藝品種。例如,某些松樹品種通過控制生長條件和授粉過程,可以培育出矮化或特殊形態的觀賞型樹木。
總之,裸子植物的有性生殖過程展現了植物界中獨特而重要的繁衍策略。這一過程既保留了某些古老的特徵,又顯示出對陸地環境的適應。深入理解裸子植物的有性生殖不僅有助於我們更好地認識植物的演化歷程,也為林業生產、生態保護和生物技術研究提供了重要的理論基礎和實踐指導。
被子植物和裸子植物是種子植物的兩大類群,它們在結構、生理和生態等方面存在顯著差異。這些差異不僅反映了兩類植物的演化歷程,也體現了它們對不同環境的適應策略。比較被子植物和裸子植物的特徵,有助於我們深入理解植物界的多樣性和複雜性。
首先,從演化時間上看,裸子植物出現得更早。裸子植物在泥盆紀末期(約3.6億年前)就已經出現,而被子植物則在白堊紀早期(約1.4億年前)才開始繁盛。這意味著裸子植物在地球上生存的時間更長,經歷了更多的環境變化和演化壓力。
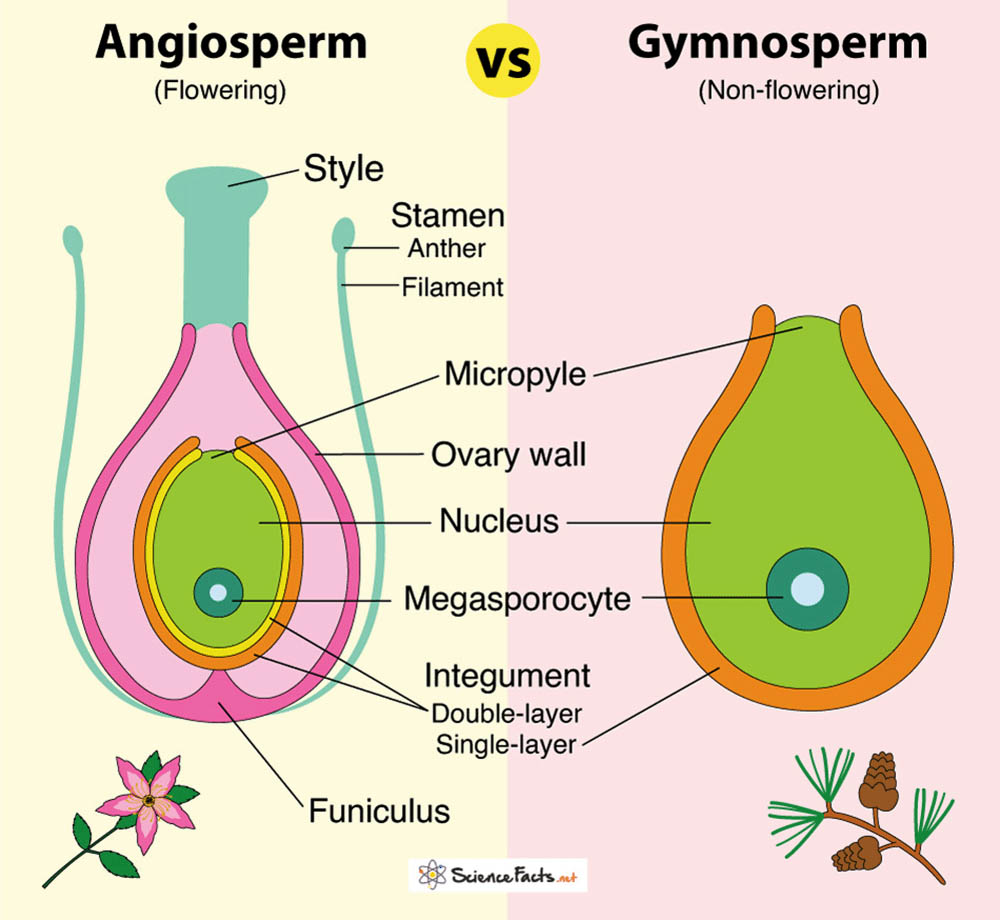
在繁殖器官方面,被子植物和裸子植物有顯著區別。被子植物的最大特徵是具有花,這是一種高度特化的繁殖器官,包括花萼、花冠、雄蕊和雌蕊。花不僅在結構上複雜,還常常具有鮮艷的顏色和香氣,這些特徵有助於吸引傳粉者。相比之下,裸子植物沒有真正的花,它們的繁殖器官是球果。球果結構相對簡單,通常由鱗片狀的孢子葉組成。
種子的形成和保護方式是兩類植物的另一個重要區別。被子植物的胚珠被包裹在子房內,受精後發育成種子,同時子房壁發育成果實。這種結構為種子提供了更好的保護,也為種子的傳播提供了多樣化的機制。而裸子植物的胚珠則直接暴露在孢子葉上,沒有被子房包裹,這就是「裸子」一詞的由來。裸子植物的種子通常只有種皮提供保護,缺乏類似果實的額外保護層。
在傳粉方式上,被子植物表現出更大的多樣性。雖然風媒傳粉在被子植物中也存在,但大多數被子植物依賴動物傳粉,如昆蟲、鳥類甚至哺乳動物。這種傳粉方式通常更加精確和高效。相比之下,裸子植物主要依賴風媒傳粉。風媒傳粉雖然簡單,但效率較低,需要產生大量花粉才能確保授粉成功。
受精過程也存在顯著差異。被子植物的一個獨特特徵是雙受精,即一個精子與卵細胞結合形成合子,另一個精子與中央細胞結合形成三倍體的胚乳核。這種機制確保了胚乳的形成,為胚胎發育提供了充足的營養。裸子植物則沒有雙受精現象,它們的胚乳是單倍體的,直接由雌配子體發育而來。
在配子體的發育方面,被子植物表現出極大的簡化。被子植物的雄配子體(花粉粒)通常只包含兩個或三個細胞,雌配子體(胚囊)通常只有七個細胞八個核。相比之下,裸子植物的配子體要複雜得多。例如,松樹的花粉粒包含多個細胞,雌配子體可能包含數千個細胞。這種差異反映了被子植物在演化過程中對繁殖結構的優化。
維管組織的結構也是兩類植物的一個重要區別。被子植物的木質部除了有管胞外,還有導管元素,這使得水分運輸更加高效。而裸子植物的木質部主要由管胞組成,缺乏導管元素。此外,被子植物的韌皮部有篩管和伴胞,而裸子植物只有篩細胞,缺乏伴胞。這些結構上的差異使得被子植物在水分和養分運輸方面更具優勢。
葉的形態和結構也存在差異。大多數被子植物的葉片較寬,具有網狀脈絡,這有利於高效的光合作用。裸子植物的葉片通常較窄,如松樹的針葉,或者像銀杏那樣的扇形葉。這種葉片形態可能是對乾旱環境的適應,有助於減少水分損失。
在生長習性方面,被子植物表現出更大的多樣性。被子植物包括草本植物、灌木和喬木,適應了各種生態位。而現存的裸子植物主要是木本植物,大多數是常綠樹種。這種差異反映了兩類植物在生態適應策略上的不同。
生活史週期的長短也是一個顯著區別。許多被子植物,特別是草本植物,可以在一個生長季內完成整個生活週期。相比之下,裸子植物通常需要更長的時間來完成生活週期。例如,許多松樹species從授粉到種子成熟需要兩到三年的時間。
在地理分佈和生態適應性方面,被子植物表現出更強的優勢。被子植物幾乎遍布全球各種生態系統,從熱帶雨林到極地苔原,從沙漠到水生環境。裸子植物雖然也有廣泛分佈,但主要集中在溫帶和亞熱帶地區,特別是在針葉林生態系統中佔據主導地位。
在物種多樣性方面,被子植物遠遠超過裸子植物。目前已知的被子植物約有25萬種,而裸子植物只有約1000種。這種巨大的差異反映了被子植物在演化過程中的爆發式輻射,也體現了它們對各種環境的適應能力。
在經濟價值方面,雖然兩類植物都對人類社會有重要貢獻,但被子植物的應用更為廣泛。大多數農作物,如水稻、小麥、玉米等,都是被子植物。此外,許多果樹、蔬菜、藥用植物和觀賞植物也主要是被子植物。裸子植物雖然種類較少,但在林業生產中占有重要地位,如松樹、杉木等都是重要的用材樹種。
在生態系統功能方面,兩類植物也有不同的作用。被子植物由於種類繁多,適應性強,在大多數陸地生態系統中扮演著關鍵的初級生產者角色。它們為多種動物提供食物和棲息地,是生態系統中能量流動和物質循環的基礎。裸子植物雖然種類較少,但在某些特定生態系統中,如北方針葉林,仍然是主要的生態工程師,對維持生態系統的結構和功能至關重要。
在應對環境stress方面,兩類植物也表現出不同的策略。許多裸子植物,特別是針葉樹,進化出了適應寒冷和乾旱環境的特徵,如疏水的角質層、耐寒的細胞結構等。這使得它們能夠在惡劣的環境中生存。被子植物則表現出更大的可塑性,能夠通過快速的形態和生理調節來應對環境變化。
在生殖策略方面,被子植物表現出更大的靈活性。許多被子植物既能進行有性生殖,也能進行無性繁殖,如通過匍匐莖、塊莖等方式。這種雙重策略增加了它們的生存和擴張能力。相比之下,裸子植物主要依賴有性生殖,無性繁殖能力相對有限。
在演化速度上,被子植物generally表現得更快。這可能與它們較短的世代時間、更頻繁的基因重組以及與傳粉者和種子傳播者的協同進化有關。這種快速演化使得被子植物能夠更迅速地適應新的環境條件。裸子植物的演化速度相對較慢,這可能是它們物種數量較少的原因之一。
在應對全球氣候變化方面,兩類植物也表現出不同的脆弱性和適應能力。一些研究表明,由於生長緩慢和遷移能力有限,某些裸子植物species可能更容易受到氣候變化的威脅。相比之下,許多被子植物由於生活史特徵和遺傳多樣性,可能具有更強的適應潛力。
總之,被子植物和裸子植物在多個方面存在顯著差異,這些差異反映了它們不同的演化歷程和生態適應策略。理解這些差異不僅有助於我們更好地認識植物界的多樣性,也為植物資源的保護和利用提供了重要的理論基礎。
植物的無性生殖是一種不涉及配子融合的繁殖方式,在植物界中廣泛存在。這種繁殖方式允許植物快速增加數量,並保持親本的遺傳特性。無性生殖在植物的自然繁衍和人工栽培中都扮演著重要角色,對於理解植物的生存策略和進行植物資源的開發利用都具有重要意義。
無性生殖的基本原理是利用體細胞分裂(有絲分裂)產生新的個體。這意味著無性生殖產生的後代在遺傳上與親本完全相同,形成所謂的克隆。這種繁殖方式的優點是能夠快速產生大量後代,並保持優良性狀。然而,它也限制了遺傳變異的產生,可能降低植物對環境變化的適應能力。
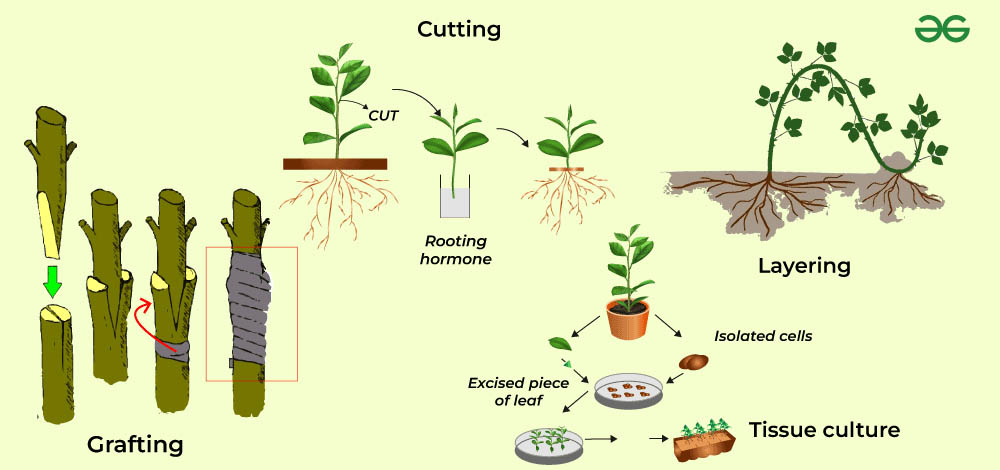
植物的無性生殖方式多種多樣,可以大致分為自然無性生殖和人工無性繁殖兩大類。自然無性生殖包括分裂生殖、營養器官繁殖和孢子繁殖等。人工無性繁殖則包括扦插、嫁接、壓條等方法,這些方法在園藝和農業生產中廣泛應用。
分裂生殖是最簡單的無性生殖方式,主要見於單細胞或簡單多細胞植物。例如,某些藻類可以通過細胞分裂直接產生新個體。裂殖酵母就是通過這種方式進行繁殖的典型例子。
營養器官繁殖是高等植物中最常見的無性生殖方式。植物可以利用根、莖、葉等營養器官形成新的個體。這種繁殖方式又可以細分為多種類型:
匍匐莖繁殖:如草莓通過匍匐莖(俗稱「草莓鬚」)在接觸土壤的節點處生根,形成新植株。
塊莖繁殖:馬鈴薯就是典型的例子,其地下莖膨大形成塊莖,上面的芽眼可以長出新植株。
球莖繁殖:如百合的鱗莖可以在鱗片基部形成小球莖,發育成新植株。
根狀莖繁殖:許多宿根花卉如鳶尾、美人蕉等通過根狀莖進行繁殖。
氣生根繁殖:一些熱帶植物如榕樹可以通過氣生根形成新的植株。
葉繁殖:秋海棠、落地生根等植物的葉片可以直接生根發芽,形成新植株。
孢子繁殖是一些低等植物和孢子植物常用的無性生殖方式。例如,蕨類植物可以通過孢子繁殖。雖然孢子是減數分裂的產物,但由於不需要配子融合就能發育成新個體,因此也被視為一種無性生殖。
人工無性繁殖方法是農業和園藝生產中廣泛應用的技術。這些方法通常能夠快速獲得大量遺傳一致的植株,對於保持優良品種的特性和大規模繁殖都有重要作用。常見的人工無性繁殖方法包括:
扦插:將植物的莖、葉或根的一部分切下,使其生根形成新植株。這是最常用的無性繁殖方法之一,適用於多種園藝植物。
嫁接:將一個植物的枝條(稱為接穗)嫁接到另一個植物的根系或莖上(稱為砧木)。這種方法常用於果樹和觀賞樹木的繁殖。
壓條:將植物的枝條彎曲壓在土中,待其生根後再與母株分離。這種方法適用於一些不易扦插的木本植物。
分株:將多年生草本植物的根系或根莖分開,每部分都能發育成新植株。這種方法常用於宿根花卉的繁殖。
組織培養:利用植物的組織或細胞在無菌條件下培養,誘導其分化形成完整植株。這是一種現代生物技術手段,可以快速大量繁殖稀有或難以用常規方法繁殖的植物。
植物選擇無性生殖作為繁殖策略有多方面的原因。首先,無性生殖可以在短時間內產生大量後代,有利於植物快速佔領有利生境。其次,無性生殖產生的後代與親本基因型相同,可以保持已經適應環境的優良性狀。此外,一些植物生活在傳粉者稀少或環境條件不利於有性生殖的地方,無性生殖成為它們的主要繁殖方式。
然而,無性生殖也存在一些局限性。最主要的問題是缺乏遺傳變異,這可能導致植物群體對環境變化的適應能力降低。此外,一些通過無性生殖產生的植物可能比有性生殖產生的植物更容易受到病蟲害的侵襲。
在自然生態系統中,許多植物同時具備有性生殖和無性生殖的能力,它們會根據環境條件和自身狀態選擇最適合的繁殖方式。例如,在資源豐富、環境穩定的情況下,植物可能更多地採用無性生殖來快速擴張種群。而在環境stress增加或資源有限的情況下,植物可能更傾向於有性生殖,以增加遺傳多樣性和適應性。
植物的無性生殖在農業生產中有廣泛應用。通過無性繁殖,可以保持作物的優良性狀,確保產量和品質的穩定。例如,許多果樹品種都是通過嫁接方式進行繁殖的,這樣可以確保果實品質的一致性。在觀賞植物培育中,無性繁殖更是不可或缺的手段,它可以保持珍稀品種的特殊性狀,如花色、花型等。
在植物保護和生物多樣性保護中,無性生殖技術也發揮著重要作用。對於瀕危植物species,可以通過組織培養等無性繁殖技術快速擴大種群數量,為物種保護贏得時間。此外,無性繁殖技術還可以用於建立植物種質資源庫,為植物遺傳資源的長期保存提供支持。
隨著生物技術的發展,植物的無性繁殖技術也在不斷創新。例如,人工種子技術將體細胞胚胎包裹在人工種皮中,模擬自然種子的結構,這種技術結合了無性繁殖的優點和種子繁殖的便利性。再如,利用基因工程技術可以在無性繁殖過程中定向改變植物的某些性狀,為作物改良提供新的途徑。
在植物進化研究中,無性生殖現象提供了有趣的研究對象。一些植物種類完全放棄了有性生殖,只通過無性方式繁衍。研究這些植物如何在缺乏遺傳重組的情況下適應環境變化,對於理解植物的進化機制具有重要意義。
無性生殖在入侵物種的擴散中也起著重要作用。許多成功的入侵植物species具有強大的無性繁殖能力,使它們能夠在新環境中快速建立種群。理解這些植物的無性繁殖特性,對於制定有效的入侵物種控制策略非常重要。
在植物生理學研究中,無性生殖過程涉及許多有趣的問題。例如,植物如何調控不同器官的再生能力,如何在脫離母體後重建維管系統等。這些問題的研究不僅有助於提高無性繁殖技術的效率,也為理解植物的生長發育機制提供了新的視角。
植物的無性生殖是一種重要的繁殖策略,在自然生態系統和人類利用植物資源的過程中都發揮著關鍵作用。深入理解植物無性生殖的機制和特點,對於植物科學研究、農業生產和生態保護都具有重要意義。隨著科技的進步,我們對植物無性生殖的認識不斷深化,相關技術也在不斷創新,這為植物資源的可持續利用和生物多樣性保護提供了新的機遇。
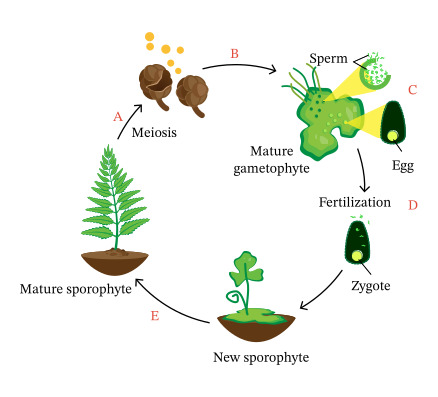
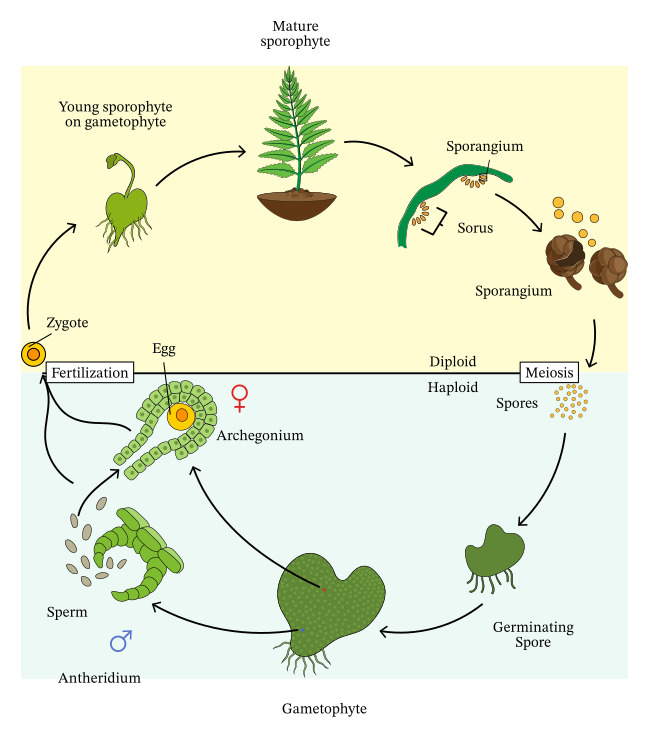
無性植物是一類主要或完全通過無性生殖方式繁衍的植物。這些植物在自然界中相對罕見,但它們的存在為我們理解植物的生存策略和進化過程提供了獨特的視角。無性植物的研究不僅在植物學領域具有重要意義,也為農業生產和生態保護提供了有價值的參考。
在自然界中,完全依賴無性生殖的植物相對較少。大多數被認為是「無性植物」的species實際上仍保留了有性生殖的能力,只是在特定環境或條件下更傾向於採用無性生殖。然而,確實存在一些plant species完全放棄了有性生殖,只通過無性方式繁衍。這些植物通常具有一些共同特徵,如繁殖器官的退化或缺失、高效的營養繁殖能力等。
蒲公英(Taraxacum)屬的某些species是無性植物的典型代表。這些蒲公英通過一種稱為「無融合生殖」(Apomixis)的方式產生種子。在這個過程中,胚胎直接從未受精的卵細胞發育而來,不需要精子的參與。這種繁殖方式使得蒲公英能夠產生遺傳上與母本完全相同的後代。無融合生殖的蒲公英在北美和歐洲的溫帶地區較為常見,它們能夠在人為干擾頻繁的環境中迅速擴張。
水燭草(Typha)是另一種常見的主要依賴無性繁殖的植物。水燭草通過地下莖(根莖)進行快速擴張,能夠在短時間內形成大面積的單一克隆群落。雖然水燭草保留了有性生殖的能力,但在許多情況下,無性繁殖是其主要的繁衍方式。這種生長策略使得水燭草在濕地生態系統中常常成為優勢species。
某些鳶尾科植物,如德國鳶尾(Iris germanica),也主要通過無性方式繁殖。這些植物通過根莖分裂形成新的植株,在園藝中被廣泛應用。雖然它們能夠開花,但種子的產生和萌發率通常很低,繁殖主要依賴於根莖的擴張。
在水生植物中,無性繁殖更為普遍。例如,水鱉(Pistia stratiotes)主要通過產生匍匐莖和子株進行繁殖。這種繁殖方式使得水鱉能夠快速覆蓋水面,在適宜的環境中甚至可能成為入侵species。同樣,浮萍(Lemna)屬植物也主要通過分裂繁殖,它們的繁殖速度非常快,在有利條件下可以在短時間內覆蓋大面積水域。
某些蕨類植物也表現出強烈的無性繁殖傾向。例如,鱗毛蕨(Nephrolepis)屬的一些species能夠通過匍匐莖快速繁殖。這些蕨類雖然能夠產生孢子,但在自然環境中,營養繁殖往往是其主要的擴張方式。
在極端環境中,一些植物完全依賴無性繁殖。例如,在南極洲的一些地區,南極髮草(Deschampsia antarctica)主要通過分蘗方式繁殖。這種生存策略使得南極髮草能夠在嚴酷的環境中維持種群。
某些沙漠植物也表現出強烈的無性繁殖傾向。例如,某些景天科植物能夠通過葉片脫落並生根的方式進行繁殖。這種策略使得這些植物能夠在水分極度匱乏的環境中生存和擴張。
無性植物的存在引發了一系列有趣的科學問題。首先,這些植物如何在缺乏遺傳變異的情況下適應環境變化?一種解釋是,這些植物可能通過表觀遺傳修飾等機制產生一定程度的表型可塑性,從而應對環境變化。另一種可能是,這些植物生活在相對穩定的環境中,不需要頻繁的遺傳變異來適應。
其次,無性植物如何避免遺傳負荷的累積?在有性生殖的植物中,有害突變可以通過重組和選擇被清除。但在無性植物中,這種機制不存在。一種假說認為,無性植物可能具有更有效的DNA修復機制,或者它們的基因組可能經過長期進化,已經高度優化,不易受到突變的不利影響。
無性植物的研究對於理解植物的進化過程具有重要意義。例如,通過比較無性植物和其近緣的有性繁殖species,我們可以了解有性生殖對植物適應性的影響。此外,研究無性植物的基因組特徵,可以幫助我們理解植物如何在缺乏重組的情況下維持基因組的穩定性。
在農業生產中,無性植物的特性被廣泛利用。許多重要的農作物品種,如香蕉、大蒜等,主要通過無性繁殖方式維持。這種繁殖方式確保了作物性狀的穩定性,有利於大規模生產。然而,這也帶來了潛在的風險,如品種單一化可能導致的病蟲害敏感性增加。
在生態保護中,無性植物的研究也具有重要意義。一些瀕危植物species可能主要依賴無性繁殖,了解這些植物的繁殖特性對於制定有效的保護策略至關重要。另一方面,一些入侵性強的植物species也可能主要通過無性方式繁衍,研究這些植物的繁殖特性有助於控制其擴散。
無性植物在環境修復中也發揮著重要作用。例如,某些能夠快速繁殖的無性植物被用於水土保持和濕地恢復。這些植物通常具有強大的繁殖能力和適應性,能夠在惡劣環境中快速建立種群。
在園藝領域,無性植物的特性被廣泛應用。許多觀賞植物品種通過無性繁殖方式維持其獨特的性狀,如特殊的花色或葉形。這種繁殖方式確保了園藝品種的一致性和穩定性。
無性植物的研究還涉及許多有趣的生理學問題。例如,這些植物如何調控其生長和發育過程?在缺乏有性生殖的情況下,這些植物是否發展出了特殊的激素調節機制?這些問題的研究不僅有助於我們理解植物生理的基本原理,也可能為作物改良提供新的思路。
在分子生物學領域,無性植物提供了研究植物基因表達和調控的獨特材料。由於這些植物的個體間遺傳背景高度一致,它們成為研究環境因素對基因表達影響的理想對象。這種研究可以幫助我們更好地理解植物如何響應環境變化。
無性植物的存在也引發了一些有關植物進化的深層次問題。例如,為什麼有些植物放棄了有性生殖?這種繁殖策略在進化上是否具有長期穩定性?這些問題的研究不僅有助於我們理解植物的演化歷程,也為探討生命的基本特性提供了新的視角。
在生態系統功能研究中,無性植物也扮演著重要角色。這些植物通常能夠快速形成大面積的單一克隆群落,對生態系統的結構和功能產生重要影響。研究這些植物如何影響養分循環、生物多樣性等生態過程,對於理解和管理生態系統具有重要意義。
總之,無性植物雖然在自然界中相對罕見,但它們的存在和特性為植物科學研究提供了豐富的素材。通過研究這些植物,我們不僅能夠加深對植物生存策略和進化過程的理解,也能為農業生產、生態保護和環境管理提供有價值的參考。隨著研究的深入,無性植物可能會為我們揭示更多關於植物生命的奧秘。

植物的無性生殖是一種不涉及配子融合的繁殖方式,可以分為自然無性生殖和人工無性繁殖兩大類。這兩種方式在自然生態系統和人類農業生產中都扮演著重要角色,對於植物的繁衍和人類利用植物資源都有深遠影響。
自然無性生殖是植物在自然條件下,不依賴人為干預而進行的繁殖方式。這種繁殖方式是植物在長期進化過程中形成的適應性策略,能夠幫助植物在特定環境下快速擴張種群。自然無性生殖的方式多種多樣,主要包括以下幾種:
分裂生殖:這是最簡單的無性生殖方式,主要見於單細胞或簡單多細胞植物。例如,某些藻類可以通過細胞分裂直接產生新個體。
孢子繁殖:許多低等植物和孢子植物通過產生孢子來進行無性繁殖。雖然孢子是減數分裂的產物,但由於不需要配子融合就能發育成新個體,因此也被視為一種無性生殖。
營養器官繁殖:這是高等植物中最常見的自然無性生殖方式,可以細分為多種類型:
匍匐莖繁殖:如草莓通過匍匐莖在接觸土壤的節點處生根,形成新植株。
塊莖繁殖:馬鈴薯的地下莖膨大形成塊莖,上面的芽眼可以長出新植株。
球莖繁殖:如百合的鱗莖可以在鱗片基部形成小球莖,發育成新植株。
根狀莖繁殖:許多宿根花卉如鳶尾、美人蕉等通過根狀莖進行繁殖。
氣生根繁殖:一些熱帶植物如榕樹可以通過氣生根形成新的植株。
葉繁殖:秋海棠、落地生根等植物的葉片可以直接生根發芽,形成新植株。
無融合生殖:這是一種特殊的無性生殖方式,植物產生的種子不經過受精過程,而是直接由未受精的卵細胞發育而來。這種方式在蒲公英等植物中比較常見。
自然無性生殖在植物的生存策略中具有重要意義。首先,它允許植物在短時間內產生大量後代,有利於快速佔領有利生境。其次,無性生殖產生的後代與親本基因型相同,可以保持已經適應環境的優良性狀。此外,在傳粉者稀少或環境條件不利於有性生殖的地方,無性生殖成為植物的主要繁殖方式。
然而,自然無性生殖也存在一些局限性。最主要的問題是缺乏遺傳變異,這可能導致植物群體對環境變化的適應能力降低。此外,通過無性生殖產生的植物群體可能更容易受到病蟲害的全面侵襲。
人工無性繁殖是人類根據植物的生物學特性,通過人為干預來實現的繁殖方式。這種方式在農業和園藝生產中廣泛應用,能夠快速獲得大量遺傳一致的植株,對於保持優良品種的特性和大規模繁殖都有重要作用。常見的人工無性繁殖方法包括:
扦插:這是最常用的無性繁殖方法之一。將植物的莖、葉或根的一部分切下,使其生根形成新植株。根據使用的植物部位,可以分為莖插、葉插和根插。不同植物適合的扦插方法可能不同,有些植物需要使用生長激素來促進生根。
嫁接:將一個植物的枝條(稱為接穗)嫁接到另一個植物的根系或莖上(稱為砧木)。這種方法常用於果樹和觀賞樹木的繁殖。嫁接可以結合不同植物的優點,如將優質品種嫁接到抗病性強的砧木上。
壓條:將植物的枝條彎曲壓在土中,待其生根後再與母株分離。這種方法適用於一些不易扦插的木本植物。壓條可以分為簡單壓條、堆土壓條、蛇行壓條等多種方式。
分株:將多年生草本植物的根系或根莖分開,每部分都能發育成新植株。這種方法常用於宿根花卉的繁殖。分株不僅可以增加植株數量,還能促進母株的更新。
組織培養:這是一種現代生物技術手段,利用植物的組織或細胞在無菌條件下培養,誘導其分化形成完整植株。組織培養可以快速大量繁殖稀有或難以用常規方法繁殖的植物,也是獲得無病毒種苗的重要方法。
人工無性繁殖在農業和園藝生產中具有多方面的優勢:
首先,它能夠保持植物的優良性狀。通過無性繁殖,可以確保後代植株與母本具有完全相同的基因型,這對於保持果樹、花卉等作物的特定品質特徵非常重要。
其次,人工無性繁殖可以大大縮短植物的繁殖週期。許多木本植物從種子到開花結果需要多年時間,而通過嫁接等方法可以顯著縮短這一過程。
第三,人工無性繁殖為難以通過種子繁殖的植物提供了解決方案。例如,一些無籽果實的品種,或者種子活力低下的植物,都可以通過無性繁殖方法進行擴繁。
第四,人工無性繁殖技術為植物的遺傳改良提供了便利。例如,通過嫁接可以將不同植物的優良性狀結合在一起,而組織培養技術則為基因工程植物的繁殖提供了重要工具。
然而,人工無性繁殖也存在一些潛在的問題:
首先,大規模的無性繁殖可能導致作物品種的遺傳單一化,增加了病蟲害爆發的風險。例如,歷史上的愛爾蘭馬鈴薯饑荒就與當時種植的馬鈴薯品種遺傳單一有關。
其次,某些無性繁殖方法可能會傳播植物病毒。例如,通過扦插或嫁接可能會將病毒從母株傳播到子代植株。
第三,長期的無性繁殖可能導致某些植物品種的退化。這種現象被稱為無性系衰退,可能與遺傳變異的積累或表觀遺傳修飾有關。
為了克服這些問題,在實際生產中,通常會將有性繁殖和無性繁殖相結合。例如,定期通過有性繁殖更新種源,然後再通過無性繁殖擴大優良個體的數量。
自然無性生殖和人工無性繁殖雖然原理相似,但在實際應用和生態意義上有顯著差異:
自然無性生殖是植物適應環境的結果,通常與植物的生存策略密切相關。例如,在資源豐富但不穩定的環境中,植物可能更多地依賴無性生殖來快速佔據有利生境。而在穩定但資源有限的環境中,植物可能更多地依賴有性生殖來增加遺傳多樣性。
相比之下,人工無性繁殖主要是為了滿足人類的需求。它的目的往往是保持特定的植物性狀,或者快速獲得大量遺傳一致的植株。因此,人工無性繁殖可能會違背植物的自然生存策略,需要人為的管理和維護。
在生態影響方面,自然無性生殖是生態系統中的正常組成部分,對維持生態平衡具有重要作用。而大規模的人工無性繁殖可能會導致農業生態系統的脆弱化,增加病蟲害爆發的風險。
自然無性生殖和人工無性繁殖都是植物繁衍的重要方式,在自然生態系統和人類社會中發揮著不同的作用。深入理解這兩種繁殖方式的特點和差異,對於植物科學研究、農業生產和生態保護都具有重要意義。隨著科技的進步,我們對植物無性生殖的認識不斷深化,相關技術也在不斷創新,這為植物資源的可持續利用和生物多樣性保護提供了新的機遇。
植物的生命期是指從種子萌發到植物死亡的整個過程。這個過程可能持續幾週、幾個月、幾年,甚至幾個世紀,取決於植物的種類和生長環境。植物的生命期不僅反映了其生理特性和生態適應策略,也對生態系統的結構和功能產生深遠影響。理解植物的生命期對於農業生產、生態保護和環境管理都具有重要意義。
根據生命週期的長短,植物通常被分為三類:一年生植物、二年生植物和多年生植物。
一年生植物在一個生長季內完成整個生命週期,從種子萌發到開花結果,再到死亡。這類植物通常生長迅速,將大部分能量投入到種子生產中。常見的一年生植物包括許多農作物,如小麥、水稻、玉米等,以及一些野花,如罌粟、向日葵等。一年生植物的生存策略是通過產生大量種子來確保下一代的繁衍。這種策略在不穩定或經常受到干擾的環境中特別有效。
二年生植物的生命週期橫跨兩個生長季。在第一年,植物主要進行營養生長,積累養分;到第二年才開花結果,完成生命週期。典型的二年生植物包括蘿蔔、胡蘿蔔、歐洲飛蓬等。這類植物在第一年通常形成葉蓮,將養分儲存在根部或其他器官中,以度過不利季節(如冬季)。第二年春天,植物利用儲存的養分快速生長,開花結果後死亡。
多年生植物的壽命超過兩年,有些甚至可以活數百年或上千年。多年生植物又可以分為草本多年生植物和木本多年生植物。草本多年生植物如牡丹、菊花等,地上部分每年死亡,但地下部分存活,來年春天再次萌發。木本多年生植物包括所有的樹木和灌木,它們的莖幹常年存在,不斷生長。多年生植物通常具有較為複雜的生命史策略,它們在生命的不同階段會調整能量分配,以平衡生長、維護和繁殖之間的關係。
植物的生命期長短受到多種因素的影響,包括遺傳特性、環境條件和生態位。從進化的角度來看,植物的生命期是對特定環境的適應結果。例如,在資源豐富但不穩定的環境中,短生命週期的策略可能更有優勢;而在穩定但資源有限的環境中,長生命週期的策略可能更為有利。
一些植物展現出極端的生命期特徵。例如,(沙漠蓍草)(Syntrichia caninervis)是一種生長在極端乾旱環境中的苔蘚植物,它可以在乾燥狀態下存活數百年,一旦遇到水分就迅速恢復生長。另一個極端是(馬達加斯加棕櫚)(Tahina spectabilis),這種植物生長數十年後只開花一次就死亡,被稱為「自殺棕櫚」。
樹木中的長壽記錄保持者是(刺果松)(Pinus longaeva),最老的個體年齡超過4800歲。這種極端長壽的特性與該樹種生長緩慢、具有高效的防禦機制有關。研究這些長壽植物不僅有助於理解植物的衰老機制,也為人類研究長壽提供了寶貴的參考。
植物的生命期與其生態功能密切相關。例如,在森林生態系統中,不同生命期的植物共同構成了複雜的群落結構。速生的一年生植物可能在林窗中迅速生長,而長壽的樹木則構成了森林的主體。這種生命期的多樣性有助於維持生態系統的穩定性和恢復力。
在農業生產中,了解作物的生命期對於制定種植策略和管理措施至關重要。例如,一年生作物通常需要密集的管理和年年播種,而多年生作物如果樹則需要長期的規劃和維護。近年來,人們越來越關注發展多年生糧食作物,如多年生小麥,以減少土壤侵蝕和提高農業的可持續性。
植物的生命期也與其繁殖策略密切相關。一年生植物通常將大量資源投入到種子生產中,以確保後代的繁衍。多年生植物則可能採取更為複雜的策略,在不同年份調整繁殖投入。例如,一些樹種會有「豐年」和「歉年」之分,在資源豐富的年份大量結果,而在資源匱乏的年份減少繁殖投入。
植物的生命期還影響其對環境變化的響應和適應能力。長壽的植物可能經歷多次環境波動,因此往往具有較強的耐受性。然而,這也意味著它們可能較難適應快速的環境變化。相比之下,短生命週期的植物可以通過快速的世代更替來適應環境變化,但可能更容易受到短期環境波動的影響。
在生態恢復和環境管理中,植物的生命期是重要的考慮因素。例如,在退化生態系統的恢復過程中,通常會先種植一些速生的先鋒物種,然後逐漸引入生命週期較長的植物species。這種策略模仿了自然演替的過程,有助於建立穩定的植物群落。
植物的生命期與其生理特性密切相關。例如,木本植物的長壽與其特殊的細胞壁結構和木質素沉積有關。這些特徵不僅提供了機械支撐,也增強了植物的抗逆性。相比之下,草本植物通常缺乏這些特徵,因此壽命較短。然而,一些草本植物通過特殊的適應性特徵,如地下儲存器官,也能達到相對較長的壽命。
植物的生命期還與其生態系統服務功能相關聯。例如,長壽的樹木在碳儲存和調節氣候方面發揮重要作用。它們能夠長期固定大量碳,對緩解全球氣候變化具有重要意義。短生命週期的植物雖然單個個體的碳儲存能力有限,但通過快速的生長和分解過程,在養分循環中扮演著重要角色。
在園藝和景觀設計中,植物的生命期是選擇species的重要考慮因素。一年生花卉可以提供季節性的色彩變化,而多年生植物和木本植物則可以提供持久的綠化效果。通過合理搭配不同生命期的植物,可以創造出既美觀又易於維護的景觀。
植物的生命期與其抗病蟲害能力也有關聯。長壽的植物通常需要發展複雜的防禦機制來抵禦長期的病蟲害威脅。這些機制可能包括物理屏障(如厚實的樹皮)和化學防禦(如產生特定的次生代謝物)。相比之下,短生命週期的植物可能更依賴快速生長和大量繁殖來應對病蟲害的威脅。
在植物進化研究中,生命期的變化是一個重要主題。例如,研究人員發現,在某些環境下,多年生植物可能演化為一年生植物,反之亦然。這種生活史特徵的轉變反映了植物對環境的適應,也為我們理解植物的演化過程提供了重要線索。
植物的生命期還與其遺傳多樣性維持有關。長壽的植物個體可能經歷多次繁殖季節,有更多機會進行基因交流,這有助於維持種群的遺傳多樣性。然而,如果種群規模小,長壽也可能導致遺傳多樣性的降低。相比之下,短生命週期的植物通過頻繁的世代更替,可能更容易產生和累積遺傳變異。
在全球變化的背景下,植物生命期的研究變得尤為重要。氣候變化可能影響植物的物候學特徵,如開花時間、生長季長度等,這些變化可能進一步影響植物的生命週期。例如,在一些地區,氣候變暖導致生長季延長,這可能有利於多年生植物,但對一些需要冷季休眠的植物species造成壓力。
植物的生命期研究對於理解和預測生態系統的動態變化具有重要意義。例如,在森林生態系統中,不同生命期的樹木構成了複雜的年齡結構,這種結構影響著森林的更新過程和對干擾的響應。通過研究植物的生命期特徵,生態學家可以更好地預測森林的長期變化趨勢。
總之,植物的生命期是一個複雜而fascinating的主題,涉及植物學、生態學、進化生物學等多個領域。深入理解植物的生命期特徵及其生態意義,不僅有助於我們更好地認識自然界的奧秘,也為農業生產、生態保護和環境管理提供了重要的理論基礎和實踐指導。
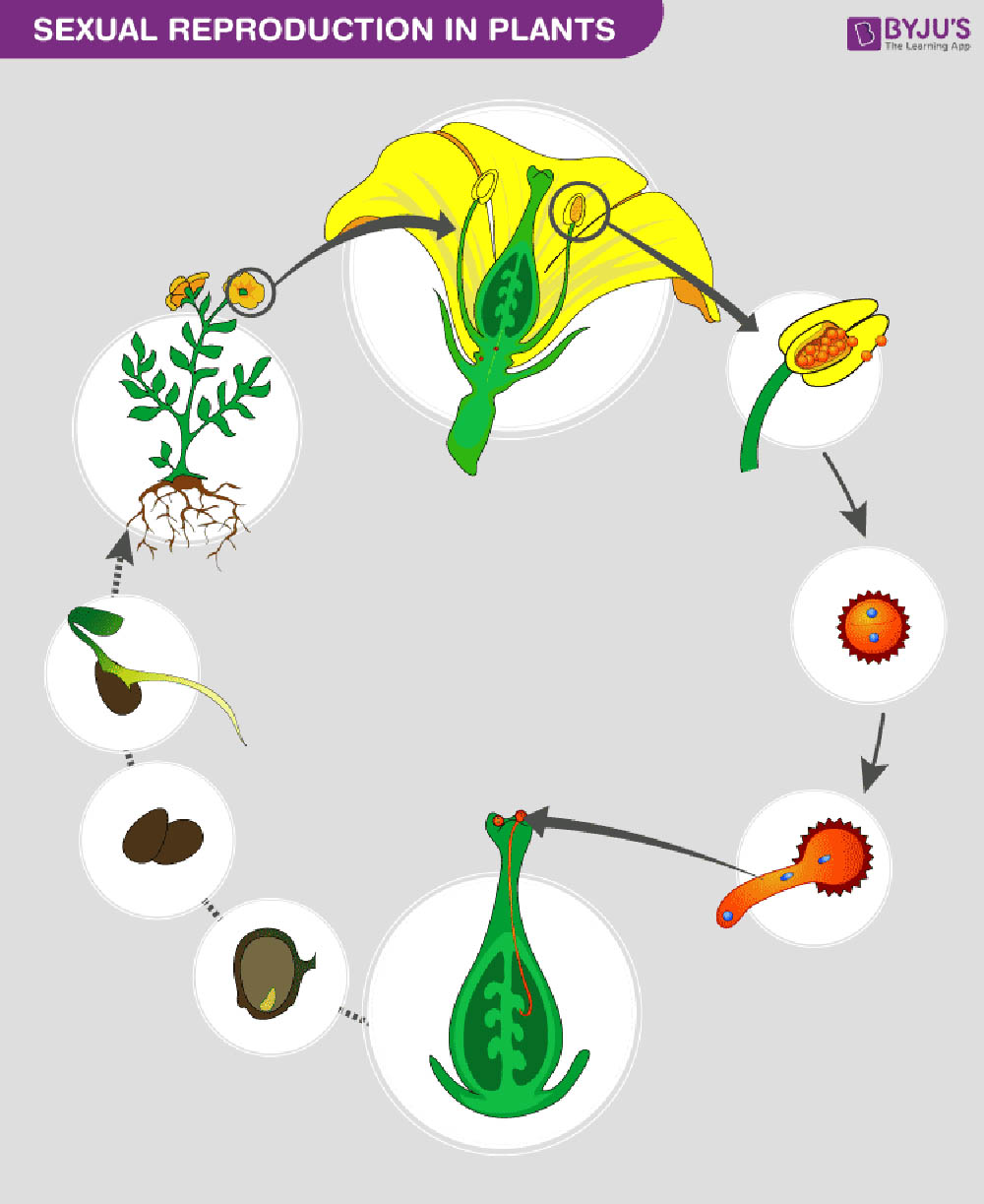
植物的有性生殖是一種涉及配子融合的繁殖方式,是植物界中普遍存在的重要生命過程。這種繁殖方式不僅確保了遺傳物質的重組和傳遞,也為植物適應不同環境提供了基礎。植物有性生殖的過程複雜而精妙,涉及多個階段和多種結構,反映了植物在長期進化過程中形成的適應性策略。
植物有性生殖的核心是配子的形成和融合。在低等植物中,如藻類,配子可能直接由體細胞分化而來。而在高等植物中,配子的形成涉及更為複雜的過程,包括減數分裂和配子體的發育。植物的雄配子通常稱為精子或精核,雌配子則稱為卵細胞。
在陸地植物的演化過程中,有性生殖系統逐漸適應了陸地環境。蕨類植物和苔蘚植物仍然依賴水分來傳播精子,而種子植物則進化出了花粉,使得精子可以通過空氣傳播,大大提高了有性生殖的效率。
被子植物的有性生殖過程尤為複雜,涉及花的發育、雄配子體(花粉粒)和雌配子體(胚囊)的形成、授粉、受精以及種子和果實的發育等多個階段。
花的發育是被子植物有性生殖的起點。花是被子植物特有的生殖器官,其發育受到複雜的基因網絡調控。花的基本結構包括花萼、花瓣、雄蕊和雌蕊。雄蕊產生花粉,而雌蕊包含胚珠,這是未來種子的前體。
雄配子體(花粉粒)的形成發生在花藥中。花藥內的小孢子母細胞經過減數分裂形成四分體,每個小孢子隨後發育成花粉粒。成熟的花粉粒通常包含兩個細胞:營養細胞和生殖細胞。營養細胞負責形成花粉管,而生殖細胞在花粉管生長過程中進一步分裂形成兩個精子細胞。
雌配子體(胚囊)的形成發生在胚珠中。胚珠內的大孢子母細胞經過減數分裂形成四個大孢子,通常只有一個存活並發育成胚囊。成熟的胚囊通常包含七個細胞八個核,這種結構被稱為(北極型胚囊)(Polygonum type)。其中最重要的是位於珠孔端的卵細胞和中央細胞。
授粉是被子植物有性生殖的關鍵步驟,指的是花粉從雄蕊轉移到雌蕊柱頭的過程。授粉方式多種多樣,包括風媒、昆蟲媒介、鳥媒和水媒等。不同的授粉方式對應著植物的不同適應性特徵。例如,風媒植物通常具有小而輕的花粉,而昆蟲傳粉的植物則可能有鮮艷的花瓣和特殊的氣味來吸引傳粉者。
授粉後,花粉在柱頭上萌發形成花粉管。花粉管的生長是一個高度精確的過程,受到多種信號分子的調控。花粉管沿著花柱生長,最終到達胚珠,將兩個精子細胞釋放到胚囊中。這一過程中,花粉管的定向生長(趨化性)確保了精子能夠準確到達目標位置。
雙受精是被子植物特有的受精方式,也是其最顯著的特徵之一。在這個過程中,兩個精子細胞分別與卵細胞和中央細胞結合。一個精子與卵細胞融合形成合子,這是未來胚胎的起源。另一個精子與中央細胞融合形成三倍體的胚乳核,這將發育成為滋養胚胎生長的胚乳組織。
受精後,合子通過一系列有絲分裂和分化最終發育成胚胎。胚胎發育的過程可以分為原胚期、球形胚期、心形胚期、魚雷形胚期等階段。在這個過程中,胚胎逐漸形成了根、莖、葉的原始結構。同時,胚乳也在快速發育,為胚胎提供營養。
種子的發育是被子植物有性生殖的重要階段。種子通常包含胚胎、胚乳(或子葉)和種皮。種皮由珠被發育而來,起保護作用。某些植物的種子還具有特殊的結構,如翅或冠毛,以助於傳播。種子發育的後期,胚胎進入休眠狀態,代謝活動降到最低水準。這種休眠機制使得種子能夠度過不利的環境條件,等待合適的時機萌發。
果實是被子植物特有的結構,由子房壁發育而來。果實的主要功能是保護種子並協助種子傳播。根據子房壁的發育方式,果實可分為真果(如蘋果)和假果(如草莓)。果實的形態和結構多種多樣,反映了植物在種子傳播策略上的多樣性適應。
裸子植物的有性生殖過程與被子植物有所不同。裸子植物沒有真正的花,而是通過球果進行繁殖。雄球果產生花粉,雌球果包含胚珠。裸子植物的受精過程較為緩慢,從授粉到受精可能需要數月甚至一年以上的時間。此外,裸子植物沒有雙受精現象,其胚乳是單倍體的,直接由雌配子體發育而來。
植物的有性生殖過程受到多種內在和外在因素的影響。植物激素在這個過程中起著重要的調控作用。例如,生長素和細胞分裂素影響花芽的形成和發育,赤黴素則參與花粉發育和果實生長。乙烯在果實成熟過程中起關鍵作用,而脫落酸則與種子休眠有關。
環境因素對植物的有性生殖也有重要影響。光週期、溫度、水分和營養狀況等都可能影響植物的開花時間和結實情況。例如,許多植物需要經過春化作用(低溫處理)才能開花,這是植物適應季節變化的一種機制。
植物的有性生殖還表現出許多有趣的適應性特徵。例如,某些植物進化出了自交不親和性機制,防止自花授粉,以增加遺傳多樣性。另一些植物則發展出了特殊的傳粉綜合徵,如(蘭科)(Orchidaceae)植物模仿特定昆蟲的外形和氣味,以實現精確的傳粉。
在農業生產中,對植物有性生殖的理解和控制至關重要。通過調控開花時間、提高授粉效率、改善果實品質等手段,可以顯著提高作物產量和品質。例如,在溫室番茄生產中,常使用蜜蜂或振動裝置輔助授粉,以提高坐果率。在果樹栽培中,了解不同品種的自交親和性對於合理搭配授粉樹至關重要。
植物的有性生殖研究為生物技術提供了重要工具。例如,通過人工控制授粉,可以實現遠緣雜交,創造新的品種。體外受精技術則為克服生殖隔離提供了可能。此外,對有性生殖過程中關鍵基因的研究,為作物的遺傳改良提供了新的靶點。
在生態學研究中,植物的有性生殖策略是理解物種分佈和種群動態的重要因素。例如,植物的開花時間和傳粉方式影響著其與傳粉者的相互作用,進而影響種群的維持和擴散。此外,種子的傳播方式和休眠特性也決定了植物對環境變化的響應能力。
植物有性生殖的研究還涉及許多前沿科學問題。例如,如何解釋被子植物的快速輻射演化?雙受精機制是如何演化而來的?這些問題的研究不僅有助於我們理解植物的演化歷史,也為探討生命的基本特性提供了新的視角。
在全球氣候變化的背景下,植物有性生殖的研究變得尤為重要。氣候變化可能影響植物的開花時間,導致植物與其傳粉者之間的時間錯配。這種現象可能對生態系統的穩定性產生深遠影響。因此,了解植物有性生殖對氣候變化的響應,對於預測和應對未來的生態變化至關重要。
總之,植物的有性生殖是一個複雜而精妙的過程,涉及多個階段和多種結構。這個過程不僅確保了遺傳物質的重組和傳遞,也為植物適應不同環境提供了基礎。深入理解植物有性生殖的機制和特點,對於基礎植物學研究、農業生產和生態保護都具有重要意義。
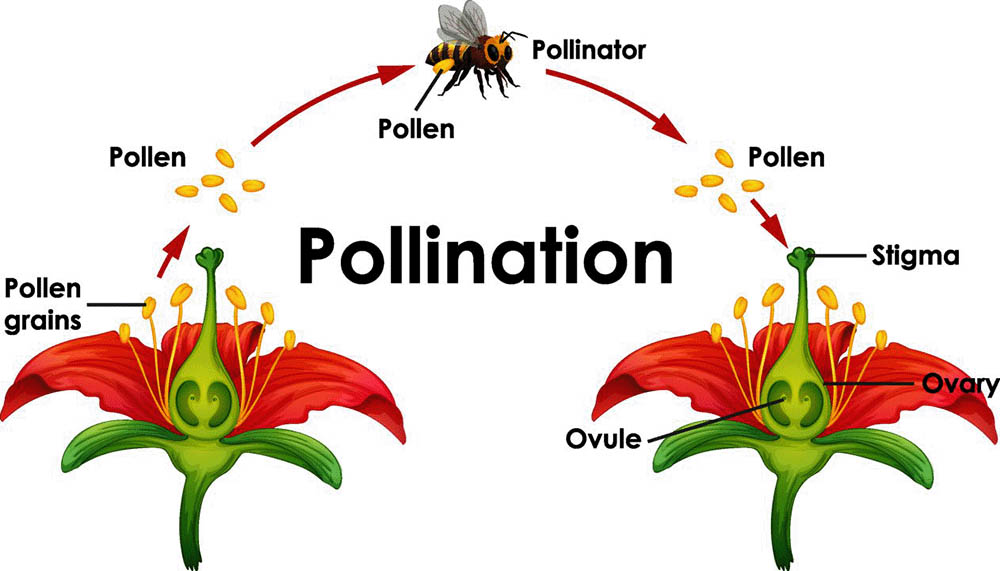
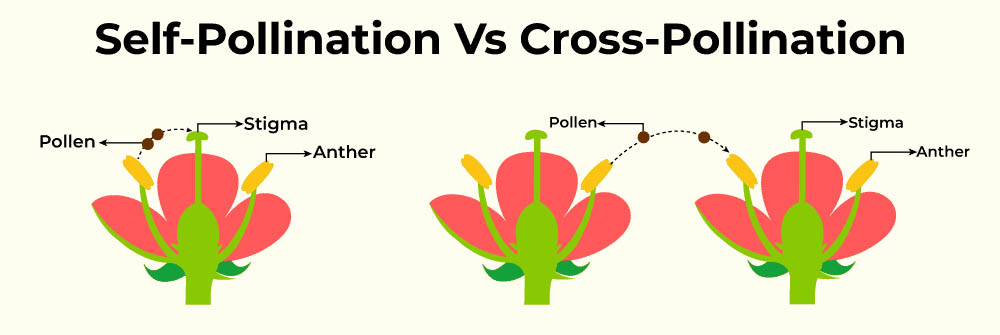
自花授粉和異花授粉是植物界中兩種主要的授粉方式,它們在植物的繁殖策略和進化過程中扮演著重要角色。這兩種授粉方式各有優缺點,反映了植物對不同環境和生態因素的適應。
自花授粉是指同一朵花的花粉落在同一朵花的柱頭上,或者同一株植物不同花朵之間的授粉。這種授粉方式可以確保植物在缺乏傳粉者或鄰近個體的情況下仍能完成繁殖。自花授粉的植物通常具有一些特殊的形態特徵,如花的結構緊湊,雄蕊和雌蕊靠得很近。
自花授粉的優點包括:
繁殖保證:即使在傳粉者稀少或環境不利的情況下,植物也能完成繁殖。
基因保存:有利於保存已適應特定環境的基因組合。
快速繁殖:有助於植物在新環境中迅速建立種群。
然而,自花授粉也存在一些缺點:
遺傳多樣性降低:長期自花授粉可能導致近交衰退,降低植物的適應能力。
有害基因累積:隱性有害基因可能在自交後表現出來。
進化潛力受限:缺乏基因重組可能限制植物對環境變化的適應能力。
許多植物進化出了防止自花授粉的機制,如雌雄異熟(雄蕊和雌蕊在不同時間成熟)、雌雄異位(雄蕊和雌蕊在空間上分離)和自交不親和性(植物能夠識別並拒絕自身花粉)等。
異花授粉是指不同植物個體之間的授粉,或者同一植物上不同花朵之間的授粉(如果是雌雄同株的植物)。這種授粉方式需要外部媒介,如風、昆蟲、鳥類等來傳播花粉。異花授粉的植物通常具有吸引傳粉者的特徵,如鮮艷的花色、特殊的氣味或者富含花蜜的結構。
異花授粉的優點包括:
增加遺傳多樣性:通過基因重組產生新的基因組合,有利於適應環境變化。
避免近交衰退:減少有害隱性基因表達的機會。
提高進化潛力:增加種群適應新環境的能力。
然而,異花授粉也存在一些潛在的缺點:
繁殖不確定性:依賴外部因素(如傳粉者)可能導致繁殖失敗的風險增加。
能量消耗:產生吸引傳粉者的結構(如花蜜、氣味)需要消耗額外的能量。
基因流失:可能導致已適應本地環境的基因組合被打破。
在自然界中,許多植物採取混合策略,既能進行自花授粉,也能進行異花授粉。這種策略使植物能夠在不同的環境條件下靈活選擇最有利的繁殖方式。
自花授粉在某些植物中非常普遍,尤其是一些自交親和的作物,如小麥、大豆等。這些植物通常具有完全花(同一朵花中既有雄蕊又有雌蕊),且雄蕊和雌蕊靠得很近。自花授粉使得這些作物能夠保持遺傳一致性,這對於農業生產來說是有利的,因為它可以確保作物性狀的穩定性。
然而,即使是主要依賴自花授粉的植物,也通常會保留一定程度的異花授粉能力。這種機制可以在需要時增加遺傳變異,有助於植物適應環境變化。例如,某些自花授粉植物在stress條件下可能會增加異花授粉的比例。
異花授粉在被子植物中非常普遍,尤其是在昆蟲傳粉的植物中。這些植物通常進化出了複雜的花結構和吸引傳粉者的機制。例如,蘭花科植物的花結構高度特化,常常與特定的傳粉昆蟲共同進化,形成了精確的傳粉機制。
風媒植物是另一類主要依賴異花授粉的植物。這些植物通常產生大量輕盈的花粉,並有特化的柱頭結構來捕捉空氣中的花粉。典型的風媒植物包括許多禾本科植物,如玉米、小麥等。
在農業生產中,了解作物的授粉方式對於作物管理和育種都非常重要。對於自花授粉作物,主要關注點是如何維持和改良純系品種。而對於異花授粉作物,則需要考慮如何確保充足的異花授粉,可能需要引入傳粉昆蟲或人工輔助授粉。
在園藝生產中,尤其是果樹栽培,了解植物的授粉需求至關重要。許多果樹需要異花授粉才能結果,因此需要合理搭配不同品種的授粉樹。例如,蘋果樹通常需要不同品種間的授粉才能獲得好的結果率。
在植物育種中,自花授粉和異花授粉都是重要的工具。自花授粉用於固定和純化特定的基因型,而異花授粉則用於創造新的基因組合。雜交育種就是利用異花授粉來結合不同親本的優良特性,創造出具有優良性狀的後代。
在生態學研究中,植物的授粉方式是理解植物-傳粉者關係和生態系統功能的重要因素。例如,異花授粉植物與其傳粉者之間的相互作用形成了複雜的生態網絡,這對維持生態系統的穩定性和生物多樣性至關重要。
氣候變化可能對植物的授粉過程產生深遠影響。例如,全球變暖可能導致植物和傳粉昆蟲的活動時間不同步,影響異花授粉植物的繁殖成功率。在這種情況下,具有自花授粉能力的植物可能具有一定優勢。
在演化生物學研究中,植物的授粉方式提供了理解物種形成和適應性進化的重要線索。例如,自花授粉可能促進物種的快速分化,而異花授粉則可能有助於維持種群的遺傳多樣性和適應潛力。
某些植物species表現出從異花授粉向自花授粉轉變的趨勢,這種現象被稱為自交進化(selfing evolution)。這種轉變可能發生在傳粉者稀少或植物分佈範圍邊緣等環境中。自交進化可能導致花的結構簡化,如花瓣縮小或消失。
另一方面,一些原本主要依賴自花授粉的植物也可能進化出促進異花授粉的機制。這種趨勢可能是為了增加遺傳多樣性或應對變化的環境條件。例如,某些自花授粉植物可能進化出更顯眼的花冠或更多的花蜜,以吸引傳粉者。
在保護生物學中,了解瀕危植物的授粉方式對於制定有效的保護策略至關重要。對於依賴特定傳粉者的異花授粉植物,保護措施需要同時考慮植物和傳粉者。而對於自花授粉植物,則可能更注重保護遺傳多樣性和防止近交衰退。
植物的授粉方式還與其繁殖系統密切相關。例如,雌雄異株的植物必須依賴異花授粉,而雌雄同株的植物則可能同時具備自花授粉和異花授粉的能力。了解這些繁殖系統的特點有助於我們更全面地理解植物的生活史策略。
在分子生物學研究中,科學家已經鑒定出了許多與植物授粉方式相關的基因。例如,與自交不親和性相關的S-基因家族,或者控制花器官發育的MADS-box基因家族。這些研究不僅深化了我們對植物繁殖生物學的理解,也為作物改良提供了新的工具。
自花授粉和異花授粉的選擇還與植物的資源分配策略有關。自花授粉通常需要較少的資源投入,而異花授粉可能需要更多資源用於產生吸引傳粉者的結構。因此,在資源有限的環境中,自花授粉可能更具優勢。
在植物地理學研究中,授粉方式的差異可能影響植物的分佈範圍和遷移能力。自花授粉植物可能更容易在新環境中建立種群,而異花授粉植物的擴散可能受到傳粉者分佈的限制。
自花授粉和異花授粉各有優缺點,反映了植物在繁殖策略上的權衡。大多數植物都能在一定程度上進行兩種授粉方式,並根據環境條件調整其比例。了解植物的授粉方式不僅對於基礎植物學研究具有重要意義,也為農業生產、生態保護和環境管理提供了重要的理論基礎和實踐指導。
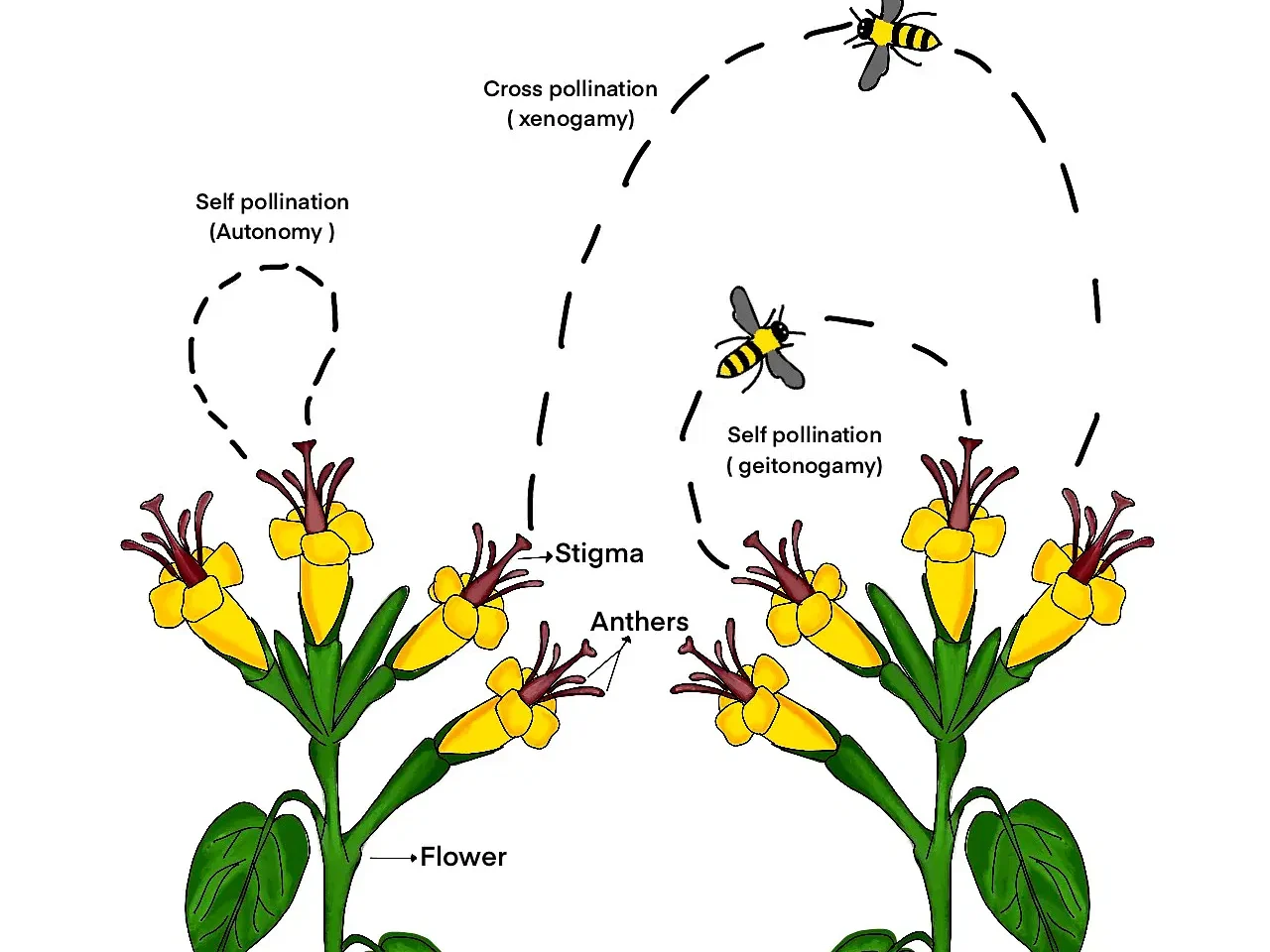
植物授粉是指花粉從雄蕊轉移到雌蕊柱頭的過程,是植物有性生殖的關鍵步驟。在長期的進化過程中,植物發展出了多種多樣的授粉方法,以適應不同的環境條件和生態因素。這些授粉方法不僅反映了植物的適應策略,也塑造了複雜的植物-傳粉者關係,對維持生態系統的平衡起著至關重要的作用。
風媒傳粉是一種常見的授粉方法,主要見於草本植物和某些木本植物。風媒植物通常具有一些共同的特徵:花小而不顯眼,通常沒有鮮艷的花瓣;產生大量輕盈的花粉;柱頭通常呈羽毛狀,有利於捕捉空氣中的花粉。典型的風媒植物包括禾本科植物(如小麥、玉米)、楊樹、松樹等。風媒傳粉的優點是不依賴動物傳粉者,可以在開闊的環境中有效傳播花粉。然而,這種方法也存在花粉浪費嚴重、授粉精確度低的缺點。
昆蟲傳粉是被子植物中最普遍的授粉方式。昆蟲傳粉的植物通常具有鮮艷的花色、特殊的氣味和富含花蜜的結構,這些特徵都是為了吸引傳粉昆蟲。常見的傳粉昆蟲包括蜜蜂、蝴蝶、飛蛾、甲蟲等。昆蟲傳粉的優點是授粉精確度高、傳播距離可以很遠。許多重要的農作物和果樹都依賴昆蟲傳粉,如蘋果、杏仁、咖啡等。某些植物與特定的傳粉昆蟲形成了高度專一的關係,如某些蘭花species只能被特定的昆蟲傳粉。
鳥類傳粉在某些地區和植物種類中也很重要。鳥媒植物通常具有管狀的花冠,富含稀薄的花蜜,花色多為紅色或橙色(這些顏色對鳥類特別有吸引力)。常見的傳粉鳥類包括蜂鳥、太陽鳥等。鳥類傳粉的一個優勢是可以在較遠的距離內進行花粉傳播,有利於增加基因交流。
蝙蝠傳粉主要見於熱帶和亞熱帶地區的某些植物。蝙蝠傳粉的花通常在夜間開放,具有強烈的氣味(對人來說可能是臭味),花冠大而堅固,能承受蝙蝠的重量。典型的蝙蝠傳粉植物包括猴麵包樹、龍舌蘭等。蝙蝠傳粉的一個顯著優勢是可以實現長距離的花粉傳播。
水媒傳粉主要見於水生植物。這些植物可能完全沉沒在水中,或者花朵漂浮在水面上。水媒植物的花粉通常能夠在水中漂浮或沉沒,以適應水中傳播。典型的水媒植物包括黑藻、眼子菜等。水媒傳粉雖然在水生環境中有效,但在整個植物界中相對罕見。
某些植物還進化出了特殊的授粉機制。例如,(無花果)(Ficus)屬植物與無花果小蜂之間形成了高度專一的共生關係。小蜂在無花果的隱頭花序內完成整個生活週期,同時完成無花果的授粉。這種關係確保了高效的授粉,但也增加了植物對特定傳粉者的依賴。
自花授粉是另一種重要的授粉方式,尤其在某些農作物中很常見。自花授粉可以發生在同一朵花內(自體授粉),也可以發生在同一植株的不同花朵之間(同株異花授粉)。自花授粉的植物通常具有完全花(同一朵花中既有雄蕊又有雌蕊),且雄蕊和雌蕊靠得很近。典型的自花授粉作物包括小麥、大豆、番茄等。自花授粉的優點是可以確保授粉成功,即使在缺乏外部傳粉者的情況下也能完成繁殖。
人工授粉是在農業和園藝生產中廣泛應用的一種授粉方法。這種方法通常用於提高授粉效率、控制雜交過程或克服自然授粉的障礙。例如,在溫室番茄生產中,常使用電動震動棒模擬昆蟲振動花朵的效果,提高授粉效率。在果樹栽培中,人工授粉可以確保授粉的及時性和有效性,提高果實的產量和品質。
某些植物species進化出了特殊的授粉機制,如爆炸授粉。這種機制在豆科的某些植物中可以觀察到,當昆蟲訪問花朵時,花瓣突然彈開,將花粉噴射到昆蟲身上。這種機制確保了花粉的精確傳播,提高了授粉效率。
偽裝授粉是另一種有趣的授粉策略。某些蘭花species的花朵模仿雌性昆蟲的外形和氣味,吸引雄性昆蟲前來"交配",從而完成授粉。這種策略雖然精確有效,但也增加了植物對特定傳粉者的依賴。
某些植物species採用欺騙性授粉策略。例如,某些(羅漢果)(Aristolochia)屬植物的花朵散發腐肉的氣味,吸引以腐肉為食的昆蟲。這些昆蟲被吸引進入花內,在尋找出路的過程中完成授粉。這種策略雖然有效,但可能降低傳粉者的再訪率。
在農業生產中,了解作物的授粉需求對於制定合理的種植和管理策略至關重要。例如,對於需要異花授粉的作物,可能需要引入授粉昆蟲或種植授粉樹。在果樹栽培中,合理搭配不同品種可以提高授粉效率和果實品質。
在生態保護中,保護植物的授粉系統同樣重要。例如,保護傳粉昆蟲的棲息地、減少農藥使用等措施都有助於維護植物-傳粉者關係。對於依賴特定傳粉者的瀕危植物species,保護措施需要同時考慮植物和傳粉者。
氣候變化可能對植物的授粉過程產生深遠影響。例如,全球變暖可能導致植物和傳粉者的活動時間不同步,影響授粉效率。此外,極端天氣事件可能直接影響傳粉者的活動和植物的開花時間。因此,研究氣候變化對植物授粉的影響對於預測和應對未來的生態變化至關重要。
在植物育種中,了解和利用不同的授粉方法是創造新品種的關鍵。例如,通過控制授粉過程,育種家可以實現特定的雜交組合,結合不同親本的優良特性。對於自花授粉作物,可能需要人工去雄和授粉來實現雜交。
植物的授粉方法還與其進化和適應密切相關。例如,某些島嶼上的植物可能因缺乏特定的傳粉者而演化出自花授粉的能力。另一方面,植物和傳粉者之間的協同進化也可能導致高度特化的授粉機制。
在分子生物學研究中,科學家已經鑒定出了許多與植物授粉相關的基因。例如,控制花香產生的基因、影響花粉管生長的基因等。這些研究不僅深化了我們對植物繁殖生物學的理解,也為作物改良提供了新的工具。
植物的授粉方法還與其生態位密切相關。例如,林下植物可能更依賴昆蟲傳粉,因為風媒傳粉在林下環境中效率較低。而開闊地帶的植物則可能更多地依賴風媒傳粉。
在生態系統服務研究中,植物的授粉被認為是一項重要的生態系統服務。許多農作物和野生植物依賴動物傳粉,這種服務的經濟價值巨大。因此,保護傳粉者和維護植物-傳粉者關係對於維持生態系統功能和人類福祉都至關重要。
某些植物species能夠根據環境條件靈活調整其授粉策略。例如,在傳粉者稀少的情況下,原本主要依賴異花授粉的植物可能增加自花授粉的比例。這種策略的靈活性有助於植物適應變化的環境條件。
總之,植物的授粉方法多種多樣,反映了植物在繁殖策略上的多樣性和適應性。了解這些授粉方法不僅有助於我們理解植物的生態和進化,也為農業生產、生態保護和環境管理提供了重要的理論基礎和實踐指導。隨著研究的深入,我們對植物授粉方法的認識不斷深化,這將為解決全球性的生態和農業問題提供新的思路和方法。
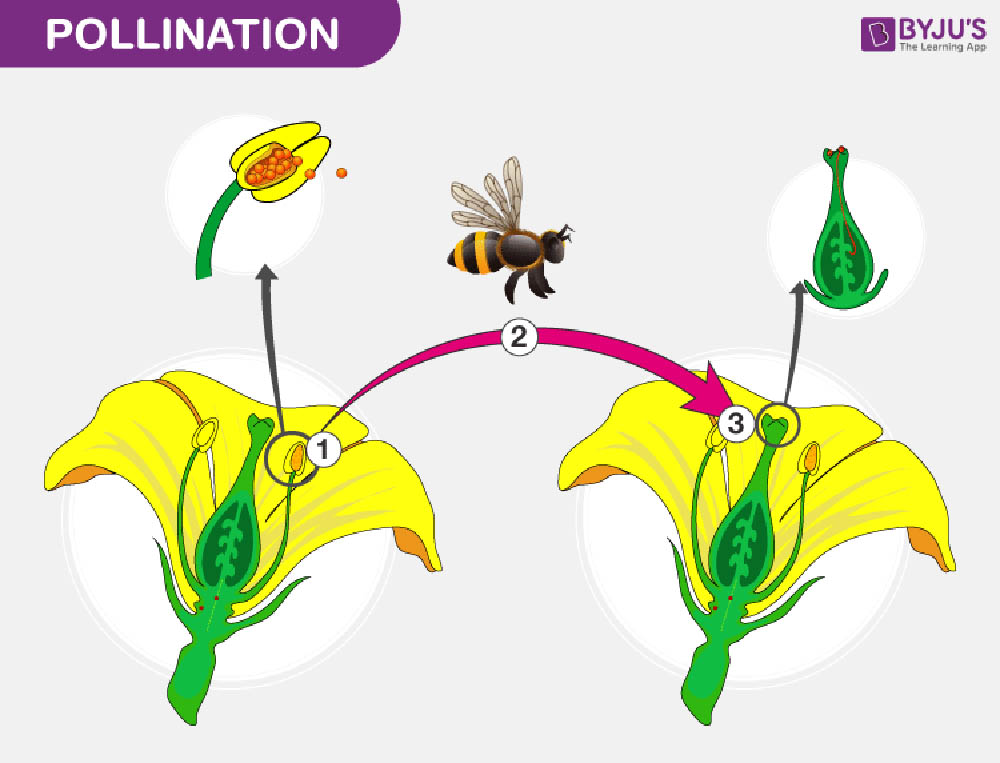
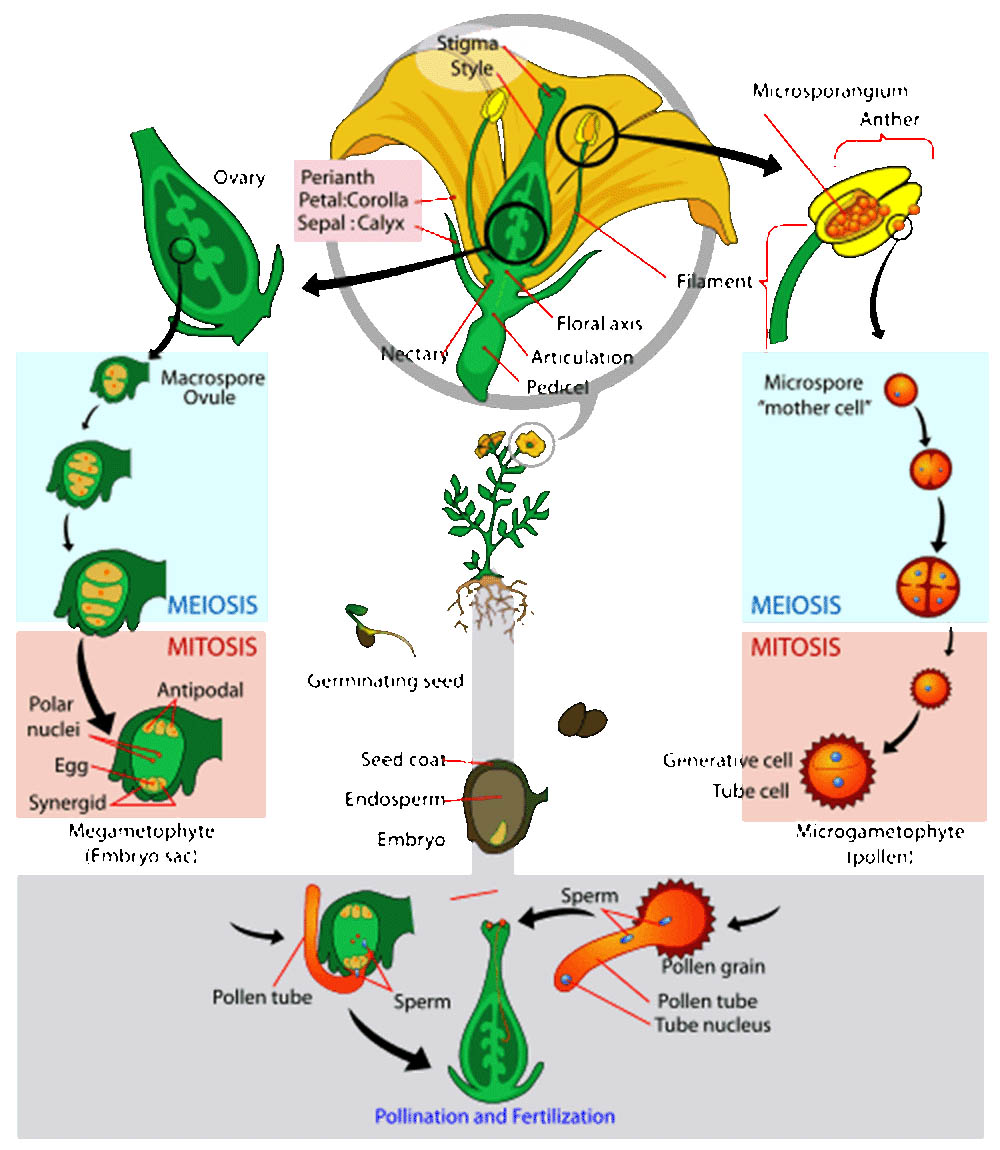
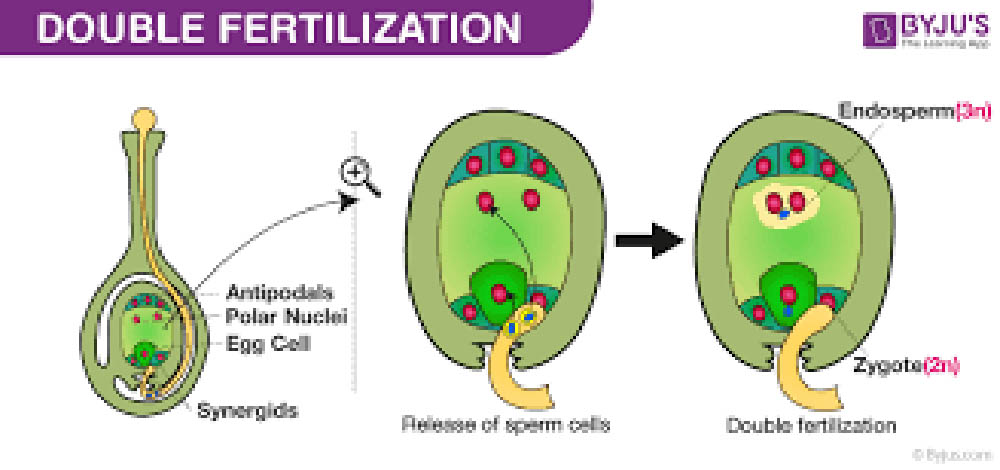
雙重受精是被子植物獨特的生殖特徵,這一過程的發現是植物學研究的重大突破之一。雙重受精不僅確保了胚胎的形成,還產生了為胚胎提供營養的胚乳組織。這一複雜而精妙的過程反映了被子植物在進化過程中對生殖效率和後代存活率的優化。
雙重受精的過程始於花粉管到達胚囊。在被子植物中,成熟的花粉粒通常包含兩個細胞:一個營養細胞和一個生殖細胞。當花粉落在柱頭上並開始萌發時,生殖細胞會分裂形成兩個精子細胞。花粉管由營養細胞形成,它會沿著花柱生長,最終到達胚囊。
胚囊是被子植物的雌配子體,通常由八個細胞組成:一個卵細胞、兩個助細胞、三個反足細胞和兩個極核。卵細胞和助細胞位於胚囊的珠孔端,而反足細胞位於胚囊的底部。兩個極核通常會融合形成一個二倍體的中央細胞。
當花粉管到達胚囊時,它會釋放兩個精子細胞。這時,雙重受精的兩個關鍵事件同時發生:
第一個精子細胞與卵細胞融合,形成合子。這個合子將發育成胚胎,是未來植物的直接前體。
第二個精子細胞與中央細胞(含有兩個極核)融合,形成三倍體的初級胚乳核。這個初級胚乳核將發育成胚乳,為胚胎的生長提供營養。
雙重受精的這兩個過程是同時進行的,這確保了胚胎和胚乳的協調發育。胚乳的形成是被子植物的一個重要特徵,它為發育中的胚胎提供豐富的營養,增加了種子的存活率和繁殖成功率。
在某些被子植物中,胚乳在種子成熟時會被完全消耗,營養物質轉移到子葉中。這種情況常見於豆科植物。而在其他植物中,如禾本科植物,成熟的種子仍保留大量的胚乳。這些差異反映了不同植物種類在種子發育和營養儲存策略上的多樣性。
雙重受精的發現對植物學研究產生了深遠的影響。它不僅解釋了被子植物種子形成的機制,還為理解被子植物的快速進化和多樣化提供了新的視角。雙重受精可能是被子植物在進化競爭中取得優勢的關鍵因素之一。
雙重受精過程的精確性和時間性受到多種因素的調控。例如,花粉管的定向生長受到化學信號的引導,確保精子細胞能夠準確到達胚囊。此外,卵細胞和中央細胞必須同時達到可受精的狀態,以確保雙重受精的順利進行。
雙重受精還涉及複雜的細胞識別和融合過程。精子細胞必須能夠識別並特異性地與卵細胞和中央細胞融合。這個過程涉及多種蛋白質和信號分子的參與,確保了受精的專一性和效率。
在分子水準上,雙重受精過程涉及多個基因的表達和調控。例如,某些基因負責控制花粉管的生長和定向,而其他基因則參與配子細胞的識別和融合。近年來的研究已經鑒定出了多個與雙重受精相關的關鍵基因,這些發現不僅深化了我們對這一過程的理解,也為作物改良提供了新的靶點。
雙重受精的結果是形成了具有三種不同基因型的組織:胚胎(合子)、胚乳和種皮。胚胎含有來自父本和母本的等量遺傳物質,是二倍體。胚乳是三倍體,包含兩份來自母本的遺傳物質和一份來自父本的遺傳物質。種皮完全由母本組織發育而來,是二倍體。這種遺傳組成的多樣性為植物提供了更大的適應潛力。
雙重受精過程中可能出現多種異常情況。例如,多精入卵可能導致胚胎發育異常。另一種情況是單受精,即只有一個精子細胞成功融合,這可能導致胚胎或胚乳發育失敗。這些異常情況在自然界中並不罕見,是植物繁殖失敗的常見原因之一。
在某些被子植物中,雙重受精過程可能出現變異。例如,在某些蘭科植物中,第二次受精(精子與中央細胞的融合)可能不會發生,而胚乳仍然能夠發育。這種情況被稱為(單受精)(Single fertilization),反映了植物繁殖策略的多樣性。
雙重受精的發現也引發了關於被子植物起源和早期進化的新思考。一些研究者認為,雙重受精可能起源於一種原始的多精入卵機制,後來演化成為現在的精確控制的過程。這個假說為解釋被子植物的快速輻射提供了新的視角。
在農業生產中,理解雙重受精過程對於作物育種和生產管理具有重要意義。例如,通過調控與雙重受精相關的基因,可能提高作物的結實率或改變種子的營養組成。此外,了解環境因素(如溫度、濕度)對雙重受精的影響,有助於制定更精準的栽培管理措施。
雙重受精還與植物的生殖隔離機制密切相關。在某些情況下,即使花粉能夠在異種植物的柱頭上萌發並形成花粉管,雙重受精也可能因物種間的不相容性而失敗。這種機制有助於維持物種的完整性,是物種形成和維持的重要因素。
在進化生物學研究中,雙重受精提供了研究基因組衝突的絕佳模型。由於胚乳包含不等比例的父母遺傳物質,它成為父母基因衝突的場所。這種衝突可能影響種子大小、養分分配等特徵,從而影響植物的適應度。
近年來,隨著顯微技術和分子生物學方法的進步,科學家們能夠更詳細地觀察和分析雙重受精的過程。例如,利用螢光標記技術,研究人員可以實時追踪精子細胞的運動和融合過程。這些新技術為深入理解雙重受精的機制提供了強有力的工具。
雙重受精還涉及表觀遺傳學的調控。例如,在胚乳發育過程中,母本和父本基因的表達受到複雜的表觀遺傳修飾的影響。這些修飾可能影響種子的大小、營養成分等重要農藝性狀。了解這些表觀遺傳調控機制對於作物改良具有重要意義。
在某些植物中,雙重受精可能導致多胚現象。多胚是指一個種子中形成多個胚胎的情況。這可能是由於多個卵細胞同時受精,或者胚胎組織的分裂所致。多胚現象在柑橘類水果中較為常見,它為研究胚胎發育和遺傳多樣性提供了有趣的研究材料。
雙重受精的研究還涉及植物生殖生物學的其他方面,如配子體的發育、花粉-柱頭相互作用等。這些過程共同構成了一個複雜的網絡,確保了被子植物繁殖的高效性和可靠性。
在生態學研究中,雙重受精過程可能受到環境因素的影響。例如,高溫或乾旱stress可能影響花粉管的生長或精子細胞的活力,從而影響雙重受精的成功率。了解這些環境因素的影響對於預測氣候變化對植物繁殖的影響具有重要意義。
雙重受精還為研究細胞融合和細胞命運決定提供了獨特的模型系統。例如,研究精子細胞如何識別並特異性地與卵細胞和中央細胞融合,可能為理解細胞識別和融合的普遍機制提供洞見。
在植物保護領域,了解雙重受精過程有助於開發新的植物保護策略。例如,某些病原體或害蟲可能干擾雙重受精過程,導致植物不育或結實率降低。通過深入研究這些干擾機制,可能開發出新的植物保護方法。
雙重受精的研究還推動了植物生殖生物學其他領域的發展。例如,它促進了人們對花粉管生長、配子體發育、種子形成等相關過程的深入研究。這些研究不僅豐富了我們對植物生命週期的理解,也為解決實際農業問題提供了新的思路。
總之,雙重受精是被子植物繁殖過程中的一個關鍵環節,它不僅確保了新一代植物的產生,還為胚胎提供了豐富的營養支持。深入理解雙重受精的機制和調控不僅有助於我們更好地認識植物的生命過程,也為農業生產和生態保護提供了重要的理論基礎。隨著研究的深入,我們對雙重受精的認識不斷深化,這將為解決全球性的糧食安全和生物多樣性保護等問題提供新的思路和方法。
種子與果實的發育是被子植物生命週期中至關重要的階段,它不僅確保了植物的繁衍,還為新一代植物提供了生存所需的初始營養和保護。這個過程涉及複雜的生理和生化變化,反映了植物在長期進化過程中形成的精妙機制。
種子的發育始於雙重受精。當花粉管將兩個精子細胞釋放到胚囊後,一個精子與卵細胞融合形成合子,另一個與中央細胞融合形成三倍體的胚乳核。合子通過一系列的細胞分裂和分化最終發育成胚胎,而胚乳核則發育成為滋養胚胎生長的胚乳組織。
胚胎的發育過程可以分為幾個階段:原胚期、球形胚期、心形胚期、魚雷形胚期等。在這個過程中,胚胎逐漸形成了根、莖、葉的原始結構。胚胎發育的模式在不同植物species之間可能有所不同,但基本過程是相似的。例如,在雙子葉植物中,胚胎通常形成兩片子葉,而在單子葉植物中,只形成一片子葉。
胚乳的發育對於種子的形成至關重要。在大多數被子植物中,胚乳是三倍體的,包含兩份來自母本的遺傳物質和一份來自父本的遺傳物質。胚乳的主要功能是為發育中的胚胎提供營養。在某些植物中,如豆科植物,胚乳在種子成熟時被完全消耗,營養物質轉移到子葉中。而在其他植物中,如禾本科植物,成熟的種子仍保留大量胚乳。
種皮是種子的外層保護結構,由珠被發育而來。種皮的發育涉及複雜的細胞分化過程,最終形成堅硬的保護層。種皮不僅保護內部的胚胎和胚乳,還可能具有特殊的結構以助於種子的傳播。例如,某些植物的種子具有翅狀結構,有助於風媒傳播。
種子發育的後期,胚胎通常進入休眠狀態,代謝活動降到最低水準。這種休眠機制使得種子能夠度過不利的環境條件,等待合適的時機萌發。休眠的調控涉及多種植物激素的作用,其中脫落酸(ABA)在維持休眠狀態中起關鍵作用,而赤黴素則在打破休眠中發揮重要作用。
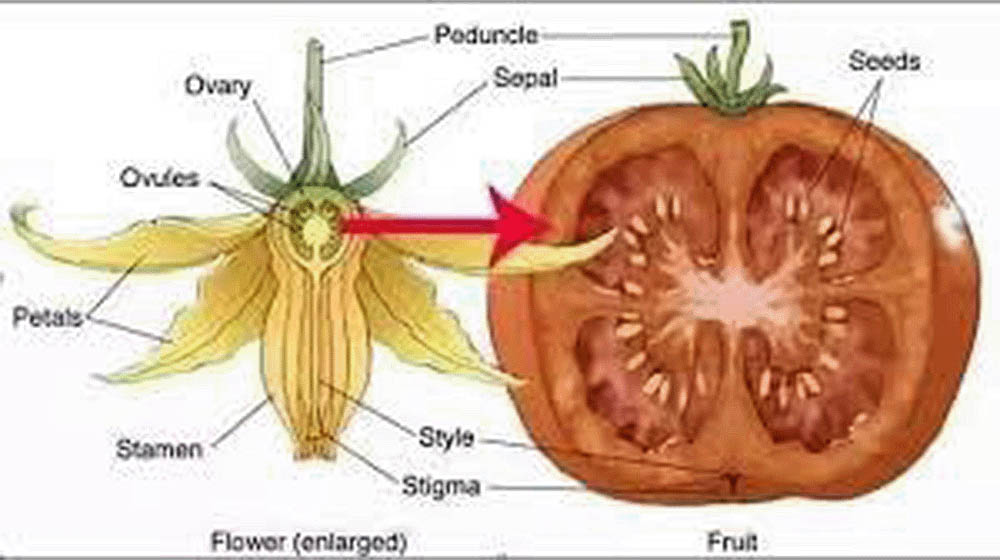
果實的發育是與種子發育同步進行的過程。果實主要由子房壁發育而來,其主要功能是保護種子並協助種子傳播。果實的發育可以分為三個主要階段:果實的設定、果實的生長和果實的成熟。
果實設定階段始於受精後子房的快速生長。這個階段涉及細胞分裂和細胞擴大,受到多種植物激素的調控,其中生長素和細胞分裂素起重要作用。在這個階段,子房壁開始分化形成果實的基本結構。
果實生長階段是果實體積和重量迅速增加的時期。這個階段主要涉及細胞擴大和細胞間隙的形成。果實生長過程中,各種營養物質不斷從植物其他部位輸送到果實中。這個階段的生長受到多種因素的影響,包括環境條件(如光照、溫度、水分)和內部因素(如激素水準、源庫關係)。
果實成熟是果實發育的最後階段,這個過程涉及一系列複雜的生理和生化變化。成熟過程中,果實通常會發生顏色變化、軟化、風味物質積累等。這些變化受到植物激素的調控,其中乙烯在啟動和協調果實成熟過程中起關鍵作用。
根據果實發育的特點,果實可以分為兩大類:真果和假果。真果完全由子房發育而來,如番茄、豆莢等。假果則包含了子房以外的其他花器官,如蘋果(花托參與形成)、草莓(花托膨大形成可食用部分)等。
果實的形態和結構多種多樣,反映了植物在種子傳播策略上的多樣性適應。例如,肉質果實(如漿果、核果)通常依靠動物傳播種子,而乾燥果實(如蒴果、瘦果)可能依靠風力或機械力傳播種子。
在農業生產中,了解和控制種子與果實的發育過程對於提高作物產量和品質至關重要。例如,通過調控授粉和受精過程可以影響種子的數量和品質。在果樹栽培中,合理的修剪、疏果等措施可以優化果實的大小和品質。
種子與果實發育的研究還涉及許多前沿科學問題。例如,如何解釋母本效應和父本效應在種子發育中的作用?胚乳的三倍體特性如何影響種子的發育?這些問題的研究不僅有助於深化我們對植物生殖生物學的理解,也為作物改良提供了新的思路。
在分子生物學研究中,科學家已經鑒定出許多與種子和果實發育相關的基因。例如,控制種子大小的基因、影響果實成熟的基因等。這些研究不僅深化了我們對發育過程的理解,也為作物遺傳改良提供了新的靶點。
種子與果實的發育還與植物的生態適應密切相關。例如,某些植物的種子具有特殊的休眠機制,使其能夠在特定的環境條件下萌發,這有助於植物適應季節性變化或不穩定的環境。同樣,果實的特性也反映了植物對特定傳播者或環境的適應。
在全球氣候變化的背景下,研究環境因素對種子與果實發育的影響變得尤為重要。例如,高溫stress可能影響花粉的活力,進而影響種子的形成。乾旱stress可能導致果實早熟或品質下降。了解這些影響有助於預測氣候變化對作物生產的影響,並開發應對策略。
種子與果實的發育還涉及複雜的代謝調控。例如,在種子發育過程中,碳水化合物、蛋白質和脂類等儲備物質的合成和積累受到精細的調控。了解這些代謝過程的調控機制,對於提高種子的營養品質和儲存性能具有重要意義。
在果實發育研究中,果實的著色過程是一個引人注目的主題。果實顏色的變化通常與特定色素(如花青素、類胡蘿蔔素)的合成和積累有關。這些色素不僅影響果實的外觀,還可能具有重要的營養價值。研究果實著色的分子機制,為培育新的果實品種提供了可能。
種子休眠是種子發育的一個重要特徵。休眠可以分為原初休眠(種子成熟時就具有的休眠)和次級休眠(種子在不利環境條件下獲得的休眠)。休眠機制的研究不僅有助於理解植物如何適應環境變化,也為解決農業生產中的問題(如種子長期保存、防止穀物穗發芽等)提供了理論基礎。
果實的後熟過程是果實發育的延續。某些果實,如香蕉、梨等,在採收後仍能繼續成熟。這個過程涉及複雜的生理和生化變化,包括呼吸跃变、乙烯產生、軟化、風味物質合成等。了解後熟過程的機制對於改善果實的儲藏和運輸品質具有重要意義。
在種子發育研究中,母本效應是一個重要課題。母本效應指的是種子性狀受到母本基因組的顯著影響。這種影響可能通過多種途徑實現,如母本組織(種皮)的直接作用、細胞質遺傳因子的影響、以及母本基因在胚乳中的偏倚表達等。了解母本效應的機制有助於更好地理解和利用種子性狀的遺傳規律。
果實的著生與脫落是果實發育研究的另一個重要方面。果實的著生涉及複雜的激素調控,主要由生長素維持。而果實的脫落則與離層的形成和活化有關,這個過程中乙烯起重要作用。在農業生產中,控制果實的脫落對於提高收穫效率和減少損失具有重要意義。
種子與果實發育的研究還與植物系統學和進化生物學密切相關。通過比較不同植物種類的種子和果實發育過程,可以推斷植物之間的進化關係。例如,被子植物特有的雙重受精和胚乳形成過程,被認為是其快速輻射演化的重要因素之一。
人種子與果實的發育是一個複雜而精妙的過程,涉及多個層面的調控和適應。深入研究這一過程不僅能夠增進我們對植物生命過程的理解,也為解決農業生產和生態保護中的實際問題提供了重要的理論基礎和實踐指導。隨著研究方法和技術的不斷進步,我們對種子與果實發育的認識將不斷深化,為植物科學和農業技術的發展開闢新的領域。
果實與種子的散播是植物生活史中的關鍵環節,它不僅確保了植物後代能夠遠離母株,減少種內競爭,還有助於植物拓展生存空間,增加種群的遺傳多樣性。植物在長期進化過程中發展出了多種多樣的散播策略,這些策略反映了植物對不同環境的適應,也塑造了複雜的植物-動物互動關係。
風力散播是一種常見且有效的散播方式,特別適合於開闊環境中的植物。風媒植物通常產生輕盈的種子或果實,具有特殊的結構以增加空氣阻力,延長飄浮時間。例如,蒲公英的種子具有冠毛結構,能夠在空中飄浮很長距離。楓樹的果實具有翅膀狀結構,可以旋轉著飄落,增加傳播距離。一些草本植物如蓍草,其整個植株在乾燥後可以被風吹走,這種現象被稱為「風滾草」。
動物散播是另一種重要的散播方式,可以分為體外散播和體內散播兩種。體外散播通常涉及果實或種子附著在動物體表。例如,某些植物的果實或種子具有鉤刺或黏性物質,可以粘附在動物的毛髮或羽毛上。牛蒡的果實就是典型的例子,其鉤狀刺可以牢固地附著在動物或人類衣服上。體內散播則主要通過動物食用果實後,將種子隨糞便排出來實現。這種方式通常涉及肉質果實,如漿果或核果。這些果實通常具有鮮艷的顏色和甜美的味道,以吸引動物食用。種子通過動物的消化道後,不僅可以傳播到較遠的地方,還可能因消化液的作用而打破休眠,促進萌發。
水力散播是水生植物和生長在水邊的陸生植物常用的散播方式。適應水力散播的種子或果實通常具有防水和浮力特性。例如,椰子的果實可以在海水中漂浮很長時間,這使得椰子能夠跨越大洋傳播。某些植物的種子具有特殊的氣室結構,增加浮力。水力散播不僅包括江河湖海中的長距離傳播,也包括雨水沖刷造成的短距離傳播。
自動散播是一些植物進化出的特殊散播機制。這種方式通常涉及果實的突然開裂或爆裂,將種子彈射出去。例如,鳳仙花的果實在成熟時會突然爆裂,將種子彈射到遠處。豆科植物的莢果在乾燥時會突然扭曲開裂,將種子拋出。這種散播方式雖然傳播距離有限,但可以確保種子離開母株一定距離。
某些植物利用重力散播,即簡單地讓成熟的果實或種子掉落。這種方式雖然看似簡單,但對於生長在陡坡或懸崖上的植物來說是很有效的散播方式。重力散播通常與其他散播方式結合,如掉落後再被動物搬運或被水沖走。
螞蟻散播是一種特殊的動物散播方式。一些植物的種子具有富含脂肪和蛋白質的附屬體,稱為油體(Elaiosome)。這種結構吸引螞蟻將整個種子搬運回巢穴。螞蟻食用油體後,通常會將種子丟棄在巢穴附近的垃圾堆中,這個環境往往適合種子萌發和生長。
某些植物利用火災作為散播的機會。這些植物的種子或果實具有耐高溫的特性,在火災後能夠存活下來。火災不僅清除了競爭者,還為這些種子創造了有利的萌發環境。例如,某些松樹的球果需要高溫才能開裂釋放種子。
人類活動也成為植物散播的重要途徑。許多植物的種子或果實可以通過人類的有意或無意行為傳播到遠處。農業活動、園藝種植、國際貿易等都可能導致植物的長距離傳播。這種傳播方式在入侵物種的擴散中尤為重要。
果實與種子的散播策略與植物的生活史特徵密切相關。一年生植物通常產生大量小型種子,採用「數量策略」以增加繁殖成功的機會。而多年生木本植物可能產生較少但營養更豐富的大型種子,採用「品質策略」以增加後代的存活率。
散播距離是評估散播效果的重要指標。不同的散播方式有不同的典型散播距離。例如,自動散播通常只能將種子傳播到幾米遠,而風力散播和動物散播可能達到數公里甚至更遠。長距離散播對於植物的地理分佈和種群基因交流特別重要。
果實與種子的散播還涉及複雜的時間動態。許多植物的果實成熟和種子釋放時間與特定的環境條件或動物行為相協調。例如,某些植物的果實在特定季節成熟,以配合傳播者的活動週期。另一些植物可能在多年中只有特定年份大量結實,這種現象被稱為「豐年結實」。
散播後的種子命運也是研究的重要方面。並非所有散播出去的種子都能成功萌發和生長。種子可能面臨被捕食、腐爛、乾枯等多種風險。因此,許多植物產生大量種子以增加成功的機會。某些植物的種子具有休眠機制,可以在不利條件下存活多年,等待合適的萌發條件。
在生態系統中,果實與種子的散播在維持生態平衡中起著重要作用。例如,許多林下植物依賴大型動物散播種子,這些動物的缺失可能導致植物群落結構的改變。同樣,某些植物種子的散播對於生態系統的恢復和重建至關重要。
氣候變化可能對果實與種子的散播產生深遠影響。例如,氣候變暖可能改變植物的物候節律,導致果實成熟時間與傳播者活動時間不同步。極端天氣事件如強風暴可能增加某些植物的長距離散播機會,但也可能對其他植物造成破壞。
在農業生產中,了解作物的種子散播特性對於田間管理和防止基因漂移都很重要。例如,某些轉基因作物的花粉或種子散播可能引起環境和食品安全方面的擔憂。因此,開發具有控制散播能力的作物品種成為一個研究熱點。
果實與種子的散播研究還涉及許多前沿科學問題。例如,如何量化和預測長距離散播事件?植物如何在散播能力和其他生活史特徵之間權衡?這些問題的研究不僅有助於深化我們對植物生態學的理解,也為解決實際問題提供了理論基礎。
在分子生物學研究中,科學家已經鑒定出一些與果實開裂和種子散播相關的基因。例如,控制果實開裂的基因、影響種子附屬結構形成的基因等。這些研究不僅深化了我們對散播機制的理解,也為作物改良提供了新的方向。
果實與種子的散播還與植物的入侵生物學密切相關。許多成功的入侵植物具有高效的散播能力,使它們能夠快速擴張到新的棲息地。了解這些植物的散播特性對於預防和控制植物入侵至關重要。
在保護生物學中,果實與種子的散播研究對於瀕危物種的保護具有重要意義。例如,了解某些珍稀植物的特殊散播需求,可能需要保護其特定的傳播者。在生態恢復項目中,選擇合適的植物種類和散播方式也是成功的關鍵因素之一。
果實與種子的散播還為生物地理學研究提供了重要線索。通過研究植物的散播能力和實際分佈情況,科學家可以推斷植物的遷移歷史和潛在的分佈範圍。這對於理解植物對過去氣候變化的響應,以及預測未來分佈變化都具有重要意義。
在進化生物學研究中,果實與種子的散播特性被視為重要的適應性狀。不同的散播策略可能導致不同的進化結果,如物種形成或滅絕。通過比較不同物種的散播特性,科學家可以推斷植物的進化歷程和適應機制。
果實與種子的散播研究還涉及複雜的植物-動物互動關係。例如,某些植物和其傳播者之間形成了高度專一的共生關係,這種關係可能是長期共同進化的結果。了解這些互動關係不僅有助於理解生態系統的複雜性,也為保護生物多樣性提供了重要依據。
