
魚類生態學 林雨莊編譯
魚類生態學 林雨莊編譯
1. 水環境與魚分類
2. 魚類生理學
3. 環境條件對魚種分佈的影響
4. 生物因數與魚類群落結構
5. 移棲地與魚群行動
6. 攝食與生長
7.生活史和種群動態
7.1 概述
7.2 生命史特徵與權衡概念
7.2.1 產卵量和卵大小
7.2.2 連續或完全繁殖
7.2.3 成熟時的年齡和大小
7.2.4 產卵後存活率
7.2.5 生殖壽命
7.2.6 親魚照顧
7.2.7 繁殖模式
7.2.8 產卵地點和時間
7.2.9 生命史理論
7.3 河流魚類的繁殖模式
7.4.1 湍急溪流的魚類
7.4.2 低地緩流的魚類
7.4 湖泊魚類的繁殖模式
7.5 河口魚類的繁殖模式
7.6 海洋中的繁殖模式
7.7.1 珊瑚礁魚類
7.7.2 大陸棚魚類的繁殖模式
7.7.3 遠海魚類繁殖模式
7.7 種群數量特徵
7.8.1 淨生殖率
7.8.2 種群生產
7.8.3 密度依賴性
7.8 魚類種群動態
7.8.1 產卵招募關係
7.8.2 種群豐度的長期變化
7.8.3 軟骨魚類的種群動態
7.9 生產
8. 魚類應用生態學
魚類在體型、棲息地和攝食方式上的多樣性與其生活史模式的多樣性相匹配(Breder和Rosen, 1966;Balon, 1975)。與攝食有關的身體形態和特徵的適應意義被解釋為與魚生活的地方和它如何生存有關。但是魚所表現的生活史模式與它所生活的環境有什麼關係呢?什麼樣的模式最有可能確保這種魚留下一些後代,從而在下一代遺傳上得到代表?
生活在相似棲息地中的成魚可能有不同的生活史特徵。一個明顯的對比是在軟骨魚類動物和硬骨類動物之間,它們共用許多遠海和底棲海洋環境。前者有內受精作用,產卵時要麼有幾個大的卵被卵箱保護,要麼生下活的幼仔。後者通常產生大量小的(直徑1-3mm)卵,這些卵在外部受精。因此,硬骨魚在受精和性成熟之間的死亡率遠遠高於軟骨魚類動物。在北海海區,80%的卵和幼體每月死亡,其中99%以上的卵未能達到幼體晚期。與硬骨魚相比,軟骨魚類動物的生活史在某些方面更接近於哺乳動物或鳥類。
即使是生活在相似棲息地中的近親物種,其生活史模式也會有所不同。在幾乎所有的太平洋大馬哈魚屬鮭魚中,成魚在繁殖一次後就會死亡,這是表面上的情況。鮭魚和鱒魚屬於近緣的鮭魚屬。一部分成魚存活下來重新繁殖。這些例子表明,生活在相似棲息地和開發相似食物資源的魚類進化出了不同的方式來確保一代中個體攜帶的基因拷貝在下一代中存在。
生命史模式不僅取決於當時和最近的生態環境,還取決於基因庫的歷史,基因庫以當時的一個群體為代表。現在的生命史必須在生態(當代)和系統發生(歷史)的約束下進行解釋。長時間的,獨立的進化歷史的彈劾和多骨魚類反映在他們不同的繁殖方式。
物種之間生活史模式差異的另一個後果可能是種群如何通過數量豐度和生物量的變化來應對環境變化。對於那些必須管理被漁業開採的種群的人來說,瞭解導致種群豐度變化的過程尤為重要。
魚能產生的卵的總體積將受到產卵前體腔中容納卵(或正在發育的胚胎)的空間的限制。卵的總體積由成熟卵的數量(繁殖力)和單個卵的體積決定。在魚類中,體型較大的魚一般不會產生較大的卵,因此繁殖力隨著體型的增大而增加(Bagenal, 1978;Wootton, 1979)。
這種關係既適用於物種內部,也適用於物種間的其他事物(表7.1)。在一個物種中,繁殖力(F)大約隨體長(L)的立方增加,即:
F = a Lb
其中a和b是參數,b大約等於4.0(圖7.1)。由於卵子的總體積是由產卵量和卵的平均體積決定的,所以在產卵量和卵子大小之間往往有一個折衷的過程(Wootton, 1984b;Elgar, 1990)。在硬骨魚和軟骨魚類動物的比較中,已經注意到數量和大小之間的這種權衡。然而,即使在硬骨船內部,這種權衡也是可以觀察到的。當性成熟時,鱈魚和鮭魚可能大小相似(1米長),但鮭魚會產生幾千個大卵(直徑4-7mm),而鱈魚會產卵幾十萬個較小的卵(直徑1.5mm)。一個簡單的計算就可以證明這些代表了相似的卵子總體積。
較大的卵產生較大的孵化後幼體。嘴的大小、遊動能力和感覺能力都會隨著體型的增大而增加,因此孵化出較大尺寸的魚能夠探測到並利用更大範圍的獵物,而且不易受到捕食。為了使進化有利於更大的卵,通過大尺寸孵化而獲得的生存收益必須補償後代數量的減少。
表7.1 英國淡水魚群的豐度、卵徑和魚長的示例
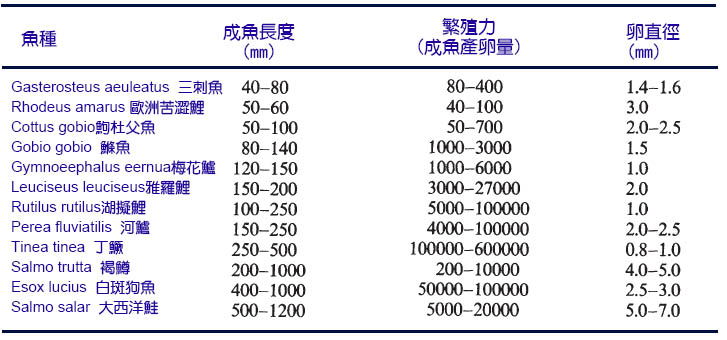
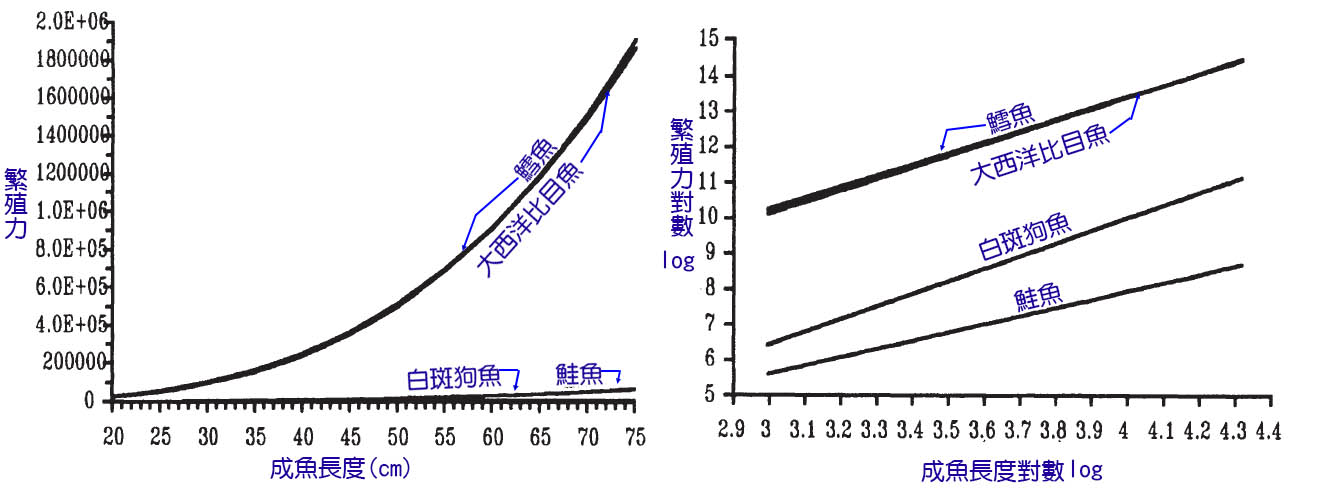
圖7.1 四個物種(鱈魚、大西洋比目魚、白斑狗魚和鮭魚的產卵量和長度之間的關係。
在一系列的產卵季節裡,一些魚的產卵量比預計的要高很多。雌性三棘背刺蜥蜴在一個繁殖季節生理上能夠產卵超過十個離合器,儘管產卵時每個離合器約占其總重量的20%。北鳳尾魚一年內產卵20次或更多。相比之下,許多物種,例如梭子魚和鱸魚,都是產卵者,在一個繁殖季節裡,幾個小時或幾天內會產生所有的卵。下一批雞蛋在一年的盛宴上不會成熟。連續產卵的一個可能的代價是,在產卵期間,魚的體型增長會減慢或停止。第二個代價可能是死亡率的增加(Pitcher和Hart, 1982年)。
生態學理論預測,在其他條件相同的情況下,在早期達到性成熟是增加後代遺傳代表性的有效途徑(Begon等人, 1989)。然而,它的代價是在較小的尺寸下成熟,因此繁殖力較低,而且可能比大尺寸成熟的魚面臨更廣泛的捕食者的風險。魚類成熟的尺寸範圍從小型鰕虎約10mm到長壽鱘和鯊魚等物種超過2000mm。在魚類中,生長率高的魚類通常會在幼年時成熟,逐漸變小,死亡率也很高(Pauly, 1980)。
在物種內部,達到成熟的平均年齡和大小因種群和環境條件而異(Alm, 1959)。一種常見但並非普遍的模式是,生長速度快的魚類成熟時年齡減少。這種減少可能伴隨也可能不伴隨著到期時規模的減少。
生物的單次繁殖和多次繁殖(Semelparity and iteroparity)是指動物物種的繁殖策略。單次繁殖的動物一生只會繁殖一次,然後死亡;而多次繁殖的動物終其一生可進行多次繁殖行為。在相似的物種中,沒有成蟲存活下來進行第二次繁殖。一個生命史意味著一些個體存活下來,一次又一次的繁殖。在iteroparous物種中,大多數大西洋鮭魚種群的存活率不到10%,再繁殖的機率很高,比如北海海區,5到15歲之間,每年的存活率約為90%。iteroparous一個缺點是每次產卵過程中投入的資源比一般物種少,因為單次繁殖(iteroparous)的成魚必須投入一些資源來保證自己的生存。通過比較semelparous和iteroparous溯河產卵鮭魚和黑鯛,結果表明,在產卵過程中,鮭形鮭魚和黑鯛使用了不到60%的身體儲備,而semelparous形式消耗了70%或更多的身體儲備(Glebe和Leggett, 1981)。
在較大的體型下達到性成熟,因而在成熟時年齡延遲的物種通常比在較早年齡達到成熟的物種具有更長的生殖壽命。這種關係在棒狀體和比目魚中都有觀察到(Roff, 1981)。其結果是,在長壽物種中,一個人將在幾個年齡段繁殖,而在任何一年中,許多年齡段的魚將在一個種群中繁殖。
軟骨魚類動物體內受精,這通常與胎生有關(Wourms等人, 1988)。在胎生形態中,約占活的軟骨魚類的65%,發育中的胚胎獲得營養的方式多種多樣。最原始的情況可能是胚胎在母體子宮內從蛋黃中獲得全部營養(卵黃餵養或卵黃營養)。在發育過程中,隨著卵黃的代謝,胚胎會失去品質。因此,胎生提供保護,防止在卵期被捕食,但不會導致比產卵更大的孵化。海底電鰩、魚雷和底棲動物棘角鯊都顯示了這種模式。Lamniformes目包括許多中上層鯊魚。在其中一些,胚胎通過吃掉其他卵子(卵子餵養或卵子吞噬)或胚胎(胚胎餵養或自噬)來獲得營養。通過這種自相殘殺,胚胎可以在出生前大大增加其品質。在長尾鯊中,每個子宮一次只有一個胚胎發育成足月。出生時,小魔豆長近1米,體重超過5公斤。(一個典型的硬骨魚的幼體孵化長度約為5mm)胚胎生長也發生在那些有母性胚胎連接的物種中,母親通過母體將營養物質傳遞給胚胎。大約27%的胎生鯊魚是胎盤性的。在這些物種中,胚胎從受精到出生的重量增加可能在6000-10000%之間。寬尾側凸鯊排卵卵直徑小於1mm,但出生時幼崽長130-150mm。沒有證據表明,在白鰭豚中,出生後,父母的照顧。
在硬骨魚類中,親代以胎生的形式照料是很少見的,儘管它已經在幾個世系中進化了。在20000多種硬骨魚中,大約有500種是胎生的。在胎生硬骨動物中,卵巢本身就是懷孕的地方,胚胎在卵巢腔或卵泡中發育。與軟骨魚類動物一樣,發育中胚胎的營養來源從依賴蛋黃到與母體的營養聯繫。在鯉科中,胎生孔雀花鱂魚guppy(Poecilia netricata:Poecilia科)在妊娠期間胚胎品質減少了25-45%,而谷鱂科的魚,胚胎品質增加了15000%。
胎生的代價是,在大多數情況下,雌性的繁殖力比相同大小的產卵物種所預期的要低得多。同樣,在增加卵子和幼年存活率以及降低繁殖力之間進行權衡。一個例外是胎生岩石魚(鮋科)(Wourms, 1991)。鮋科魚類表現出一系列的情況,從典型的硬骨魚模式產卵大量小的遠海卵到胎生。這是在一個亞科中進化而來的,其中包括平鮋屬(Sebastes)岩石魚。受精是內部的,卵子在卵巢腔內發育。然而,這些胎生石魚的繁殖力並沒有顯著降低。幼體雖然器官系統發育良好,但出生時的長度不到10mm。在懷孕期間,發育中的胚胎可以從死卵和胚胎的分解中獲得一些營養,也可以從它們自己的卵黃中獲得一些營養。懷孕的石魚必須提供數千個發育中的胚胎所需的氧氣,並處理它們的廢物。
在產卵(卵生)物種中,父母的照顧可能僅限於在產卵前準備產卵地點。七鰓鰻用身體移動石頭或用吸吮嘴把石頭運走,以此來挖窩坑。產卵後卵被石頭覆蓋。在溪流產卵的鮭魚中也可以看到類似的模式。一旦卵被覆蓋,就不會有進一步的父母照顧。
受精後對卵和幼體的照顧發生在22%的硬骨魚科中,儘管不是一個科的所有物種都必須如此(Sargent和Gross, 1986)。最常見的形式是父親照顧(表7.2)。通常情況下,父母的照顧是在守衛,而父母則保護卵不受掠食者的傷害。在太陽魚、刺魚和一些瀨魚的例子中,雄性築巢產卵,然後加以保護。產卵在基底上或巢中,有耗盡周圍水的氧氣或被沉澱物掩埋的危險。卵的通風通常伴隨著護卵,親魚在卵上漂浮著水流。有幾個科,例如蟬科,包括父母在嘴裡孵卵的物種,而在知更鳥中通常是母親。父母輕輕地攪動雞蛋,使它們保持通風。即使在孵化之後,這些幼小的知更鳥仍然離它們的母親很近,如果有危險,它們會飛回媽媽的嘴裡。在一些硬骨魚中,卵被帶到母魚的表面(Balon, 1975)。在管魚和海馬中,雌魚將卵沉積在雄性魚腹部的凹槽或育兒袋中。
當雄性魚來照看卵子時,雌性的繁殖力就不必減少了,作為父母照顧的交換條件。父母的雄性很可能會付出增加死亡率和降低生長速度的代價。在那些有女性或雙親護理的情況下,生育能力會降低。例如,在雙卵圓線蟲中,如果雌性照顧自己的卵和幼仔,連續產卵的間隔要比受精後立即取卵時長得多。
表7.2 以魚科(家庭)百分比表示的魚類父母照顧分佈。Sargent and Gross (1986)
親魚照顧模式 軟骨魚 硬骨魚
雄魚照顧卵 6 11
雌魚照顧卵 66 7
無照顧 28 78
----------------------------------------------------------------------
大多數魚類具有不同的性別-雌雄異體性-內部或外部受精。而軟骨魚綱之板鰓亞綱(鯊魚、鰩魚等)內部受精;而大多數硬骨魚類外部受精。 但也有其他性行為繁殖方式。
在脊椎動物中,一個個體既可以作為雌性又可以作為雄性發揮作用的情況很少見,但在硬骨魚類中也發現了一些例子(Warner, 1978)。在同時性或同步性兩性畸形中,個體同時具有功能性卵巢和睾丸,因此自我受精是可能的。幾種深海魚類同時是雌雄同體,比如鱸形科(Serranidae),生活在亞熱帶和熱帶淺水區。在北美洲東南部的沿海沼澤中也發現了一種小型鯉科動物Rivulus,它是一種自我受精的雌雄同體。在連續的兩性體中,個體首先具有一種性別的功能性性腺,然後在生命史的某個時刻,性腺經歷了一個轉變,變成了另一個性別的功能。在原雄蕊形態中,男性有功能睾丸的階段先於有功能卵巢的女性階段。在原雌雄同體的物種中,雌性階段先於雄性。在一些改變性別的物種中,可能有個體,主要的雄性或雌性,不會改變性別。在這些性別變化的連續兩性體中,研究得最好的是礁棲魚類,包括瀨魚(唇形科)、鸚嘴魚(鸚哥魚亞科)和鰕虎(鰕虎科)(Warner 1978年)。
台灣養殖石斑魚研究也發現溫度與性別轉變之關係。石斑魚的體型大小是決定雄性繁殖能力的重要因素,大型的雄魚由於擁有比較大的地盤和比較發達的生殖腺,要比小型的雄魚有更多的機會參與交配。 因此,演化使得每一個個體在體型小、不適合扮演雄魚角色時,都先成為雌魚,以避開繁殖上的劣勢。
目前有許多可能的解釋,例如在自然情況下魚類會性轉變是由於族群因素,也就是在同族群中,本來雄魚與雌魚的比例構成一個穩定的雄雌關係,若是把雄的移走,那其中一尾雌魚的會在數天內發育成為雄魚的。另外一種可能是,由於性轉變可以獲得更有利的生殖策略,因此年紀較小時(較小體型)是雄魚的,年紀較大且體型較大時變為雌魚的,例如黑鯛,因為較大體型的雌魚產卵個數較多,有利於族群存活。也有一種可能是避免「近親交配」,黑鯛長到第三年才變成雌魚,交配對象是出生一、二年不同親代產生的成魚,因此可避免近親交配的現象。這種種可能的原因促使雌雄同體的魚類,在長期演化的過程中,慢慢發展出具有特色的性轉變生殖策略。
所有只有雌魚基因代代遺傳的雌性種群在脊椎動物中也很少見。在北美洲西南部乾旱地區的小泉和溪流中,有兩個屬的鯉魚發生孤雌生殖(Poecilia和Poeciliopsis)(Vrijebhoek, 1984)。這些單性生殖的形式很奇怪,因為它們是共存和密切相關的有性生殖物種的精子寄生蟲。為了使孤雌生殖形式的卵子發育,它們必須被有性生殖物種的雄性精子啟動。在一種模式中,父系基因決定了後代的基因型,但只有母體基因一代一代地遺傳下來,這就是所謂的雜交發生系統。在另一個系統,雌核發育,精子只啟動卵子。父系基因沒有被納入發育中胚胎的基因組中。
繁殖成功與否也取決於棲息地的位置和一年中何時產卵。這些特徵最好是在繁殖的棲息地中考慮的。
生命史理論試圖預測在特定的環境條件下可能進化的特徵組合(Begon等人, 1989)。從目前的形式來看,這一理論表明,生命週期中生殖前和生殖階段的相對死亡率是重要的。如果生殖前死亡率高且不可預測,則有利於延遲生殖、延長成年壽命和比例。如果成人死亡率很高且不可預測,那麼早期生殖和對早期生殖的高投資是有利的。
在低級序溪流中,高流速和可變流速對產卵魚類既有利又不利。其優點包括高含氧量的水,其流速可防止沉積物在卵上堆積。一個主要的缺點是,魚卵和幼魚有被沖到下游並被殺死的風險。
在北美和歐亞大陸的冷水、旱地溪流中,最常見的物種是鮭科、杜父魚科和一些鯉屬。鮭魚把卵埋在礫石巢穴裡,那裡有很好的水流。產卵時,巢穴周圍的區域會受到有力的保護,尤其是雄魚。許多雄性鮭魚會發展出第二性特徵,即長有尖牙的鉤狀下顎,用來與其他雄性進行搏鬥。在杜父魚科中,雄魚為石頭之間或下方形成的洞辯護。雌性在洞裡產卵,卵附著在牆上。雄魚給卵通風,保護卵不受入侵者的侵襲。溪流上游的鯉魚,如歐洲的小鯉魚,把卵撒在岩石和石頭之間的縫隙裡,但不守衛這個地點。在北美東部,許多小型鯉魚守護著卵(Moyle和Herbold, 1987)。
溪流產卵鮭魚的卵是硬骨魚產生的最大的卵之一,其繁殖力也相應較低(表7.1),根據體型大小有幾百到幾千個。溫帶海洋和淡水物種的卵直徑比較表明,淡水物種(包括河流和湖泊物種)的卵往往較大(圖7.2)(Wootton, 1979;Elgar, 1990)。從大卵孵化出來的較大的幼體在應對水流和開發更大的獵物方面可能有優勢。在自來水中,浮游植物和小型浮游動物通常以低密度存在。
長期生活在這些低階河流中的魚類通常體型較小。它們通常是連環產卵的。大型魚類通過向上游遷徙來開發產卵的棲息地。例如溯河產卵的七鰓鰻和鮭魚,它們是全部的產卵者,而且在許多情況下都是類似的。
鮭魚在秋季或冬季產卵,卵埋在礫石中發育緩慢。即使在孵化之後,這些被稱為alevins的幼鳥在耗盡卵黃儲備時仍留在礫石中。鮭魚魚苗只在春季從沙礫中冒出,硬骨魚的體型大約為30mm。相比之下,這些低階溪流中的棉類、鯉類和泥狗魚類在春季或夏季產卵。普遍較高的溫度意味著卵孵化只需幾天時間,而幼仔出來的長度不到10mm。
低緯度低階河流的一個特徵是胎生鯉魚的存在。例如,在中美洲和南美洲的小溪流中生活著幾種花鱂魚。對千里達Trinidad河裡的古比河的實驗和實地研究支持了生命史理論的預測(Reznick等人, 1990年)。在食魚動物主要以成年古比魚為食的地方,與以幼魚為主的地方相比,食魚魚魚成熟的年齡要早一些,並且產下的幼魚數量也越來越小。
在美國西南部和墨西哥乾旱區的溪流中,生活著孤雌生殖的花鱂魚(Poecilia、Poeciliopsis)。在這種環境中出現這種不尋常的繁殖方式的一種解釋是”凍結生態棲位”假說(Vrijenhoek等人, 1987年)。這表明,單性生殖是一種減少種內競爭的機制,在這種環境中,其他魚類種類很少,種間競爭也很低。所有・雌性克隆體,每一個都由基因相同的個體組成,但在基因上與其他克隆體不同,它們都來自於一個遺傳變異的性祖先。克隆間的選擇產生了一種組合,這種組合以減少克隆之間生態重疊的方式劃分食物和空間。
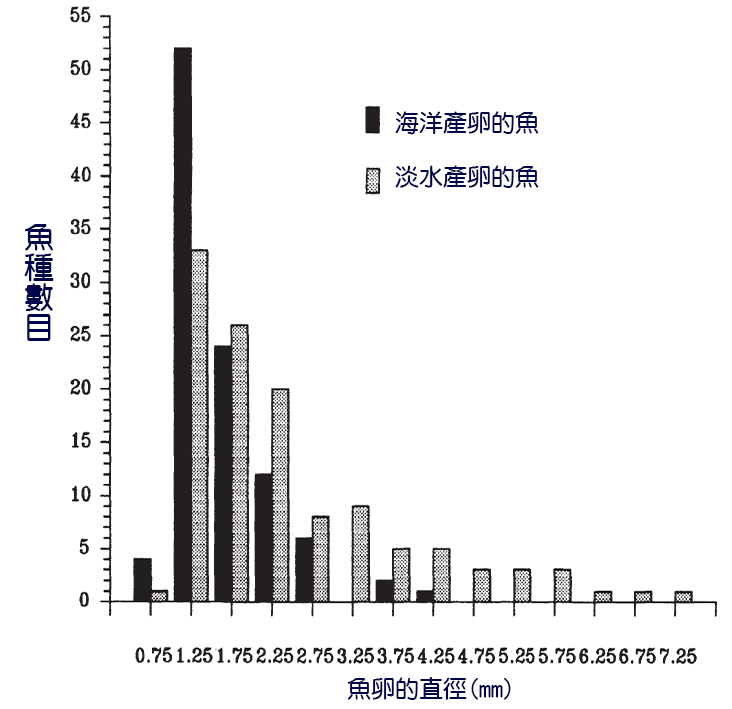
圖7.2 北溫帶海洋水域和淡水水域魚類的卵直徑。
對許多低緩河流河的魚類來說,它們的繁殖生物學適應了洪水的周期。產卵通常發生在枯水和洪峰之間的某個時間段,只有在洪水下降時才很少發生(Welcomme, 1985)。這個時間安排確保幼仔孵化到一個覓食棲息地面積不斷擴大的環境中。在高緯度地區,產卵時間也與季節性氣溫升高和春夏提前有關。這一時期通常與積雪融化時的高水位相對應。
整批同時期產卵在低緩河流魚類中很常見,這可能是因為洪水週期中有一段時間對卵的發育和幼魚的生長最為有利。在熱帶河流中,例如許多較大的南美輪藻和非洲鯉魚。在溫帶地區,鯉魚和其他鯉魚、梭子魚和鱸魚都是全部的產卵者。連環產卵器也出現了。它們通常是壽命短的小魚。大多數的低緩河流魚類都是多次繁殖(iteroparous)。
產卵地點可以在主河道或淹沒的植被中。在亞馬遜河的一條支流馬德拉河中,大型特徵魚遷徙到主河道產卵,然後移到被洪水淹沒的森林底部覓食(Goulding, 1980)。年輕人被沖到下游的靜水和氾濫的平原湖泊。在中國的河流中,大型鯉魚,如鱅魚,在河中產卵;這些卵被帶到下游,並沿著被洪水淹沒的邊緣進入育雛區。水的渾濁使卵和幼體免受捕食,而水的運動也使卵保持充氧和無沉積物。
另外兩個重要的產卵地點是植物叢和石頭堆的基質(Balon, 1975)。像梭子魚、鯉魚和藤壺這樣的物種會把卵附著在水下的水生植物上。其他一些物種,如玻璃梭鱸、吸盤魚,萊茵鯉魚屬,把它們的卵撒在岩石和石頭上。還有一些像湖擬鯉,鯛魚和鱸魚,把它們的卵附著在沉入水中的植物、原木或石頭上。
無脊椎動物或脊椎動物捕食卵和幼體的風險在低緩河流魚類中比在陡急溪流中更高。在所有的產卵者中,短時間內產生大量的卵,這可能會淹沒捕食者。其他的魚守護著它們的卵。在像脂鯉這樣的物種中,被保護的卵附著在植物上。巨骨舌魚在洪水淹沒的地面上形成了一個空洞。東南亞的雄性攀鱸(anabantoids)製造一個漂浮的泡沫巢來保存卵。這就保證了卵在靜止、溫水中接近富氧表面膜,容易發生去氧。在溫帶水域,雄性刺蝟在水底或植被中築巢。雌性苦口魚,鰟鮍(Rhodeus amarus),通過一個長的管狀產卵器將卵沉積在淡水貽貝的外套腔中。吳郭魚會在感潮的淺水泥地上用尾鰭掃出一個個淺圓盤,然後產卵魚其中,減少被其他魚類捕食的機率。
大多數河漫灘物種避免了產卵時由於水位下降而暴露在外的危險。然而,南美洲和非洲的一些鯉科魚類可以在定期乾涸的池塘中維持種群數量(Wourms, 1972)。這些短壽命的“一年生”小魚產卵,當池塘排空時,它們可以在底泥或植被中存活。卵進入滯育期,在此期間卵發育受阻。當池塘重新注滿時,進一步的開發就得到了刺激。
湖泊中的沿岸魚類具有許多河漫灘魚類的繁殖模式。事實上,一些湖泊魚類遷移到河流中產卵,利用了洪水造成的環境(Lowe-McConnell, 1987)。在這些物種和溫帶湖泊的沿岸物種中,產卵是季節性的。
溫帶湖泊魚類的主要產卵期為春季至盛夏。有一些明顯的例外。江鱈在隆冬產卵。湖鱒魚在秋天產卵,而許多白魚也在秋天或冬天產卵。卵是在礫石上產卵的,但是,對於江鱈和一些白魚來說,它們的卵稍有浮力,因此在水的運動下會輕輕地移動。
在低緯度的湖泊中,一些物種有不同的產卵季節,而其他物種全年產 卵。即使是後一種物種,也往往有一個時期,種群處於繁殖狀態的比例也比其他時期高。熱帶湖泊季節性產卵的優勢在於溫度和日長只有微小變化,在某些物種中, 這可能反映出浮游生物產量的變化,這與乾濕季節性或風驅動周期模式有關。在尼加拉瓜的吉洛亞湖,在沿海地區產卵的慈鯛科魚類呈現出季節性繁殖,這可能反映出對食物供應和產卵地點競爭的反應。在雨季,優勢物種厚唇雙冠麗魚 (Cichlasoma citrinelum)佔據了大部分可用的產卵場地。在旱季,當其他慈鯛科魚類能夠產卵時,它們停止繁殖(McKaye, 1977)。
湖泊物種包括全部和連續的產卵者,對於沿海物種,某種形式的親魚照顧是常見的。像藍鰓太陽魚這樣的中央生殖道的雄性會清理底部雌性產卵的巢穴。在藍鰓太陽魚中,雄性有一個包括替代生殖策略的繁殖系統(Gross, 1984)。有些雄性在6或7歲時成熟,築巢並在裡面保護卵。其他雄性在2歲時成熟,體型較小。這些雄鳥並不試圖築巢,而是潛伏在大型親代雄性的巢穴周圍,在親代雄性與雌性產卵時,沖進來讓一些卵受精。隨著它們變大,這些“偷偷摸摸的”雄性開始變得像雌性,這使得它們能夠接近父母雄性的巢穴並偷偷受精。包括一些溯河產卵鮭魚在內的其他魚類也發現了涉及大型“領地”和小型“偷襲”雄性的替代交配策略(Gross, 1984)。
1972年,許多雛魚棲息在非洲湖中。雄魚在底部築巢,在某些物種中,巢是精心建造的。這些巢穴聚集在一起,雄魚在巢穴周圍保衛小領地。產卵後,雌魚叼起嘴裡的受精卵,離開雄魚孵化卵和幼魚。
雖然河口被許多海洋魚種用作育雛區,但只有少數物種在那裡繁殖。這種物種往往是體型小,連續產卵和某種形式的親魚照顧是常見的。例如鰕虎和棘背魚科。少數物種利用潮汐週期來降低其卵被水生食肉動物捕食的風險。這些物種在漲潮時產卵,將卵產在潮濕的植被或潮濕的沙灘上。隨著潮水退去,這些卵雖然仍處於潮濕的環境中,但還是冒了出來。孵化發生在隨後的高潮時,此時卵再次浸入水中。由於潮汐週期和月相的關係,這些物種通常有一個潮汐產卵週期。大西洋銀漢魚、美洲原銀漢魚,產卵在滿月或新月潮差最大的時候。
大西洋銀漢魚有一個相對不尋常的性別決定機制(Conover和Heins, 1987)。在大多數脊椎動物中,個體的性別是由基因型決定的。在一些美洲原銀漢魚種 群中,性別是由性別決定基因和環境溫度之間的相互作用決定的:環境性別決定的一個例子。在繁殖季節開始時水溫較低時,大多數後代都會變成雌性。在這個季節 結束時,在高水溫下產生的後代會變成雄性。這種效應是適應性的,因為雌性的生長期較長,因此體型比雄性大。女性的生殖力隨著體型的增大而增加,而男性美洲原銀漢魚則沒有證據表明生育能力隨體型的增大而增加。在繁殖季節較短、溫度不能很好地反映時間的北方種群中,這種環境對性別決定的影響很弱或根本不存在。
珊瑚礁魚類的種數很難界定,因為如果把礁區附近夾雜的沙地、海草床乃至中表水層的魚類,只要是暫時游經、季節性棲息或偶爾游入之大洋性洄游魚類,都予計入,則種數可達一萬種,占所有魚類的30%左右。臺灣約有1500種珊瑚礁魚類。
珊瑚礁魚類的多樣性之高,令人驚歎,如一塊5x5平方米之礁塊可紀錄高達上百種的魚類在此共存。這主要歸功於魚類與無脊椎動物及海藻間長期共同演化的結果,以及資源分配說(resources partitioning),亦即牠們在棲所、食性及日夜時間上之資源均各有不同,使得競爭減少;彩票學說(lottery theory)是指同一個棲所一但騰空,誰先到誰先贏,因此種類就不固定,種數會較多。
淺海珊瑚礁只占地球表面積的0.2%-0.3%,分布在南北緯30度間之海域,水深在50公尺內之溫暖、清澈、陽光充足的海域內。由於順時針的北太平洋環流和北大西洋環流流徑赤道,把暖水帶到了珊瑚礁全球分佈之兩大熱點區-東印度群島及加勒比海。前者的種數為後者的6倍。
珊瑚礁魚類不但色彩豐富艷麗、模樣可愛,更有許多有趣的食衣住行的生活習性,或是令人讚嘆的共生、擬態、掠食、競爭的行為;多樣的生殖方式等。
珊瑚礁環境逐漸劣化,不利於生物多樣性。許多過去屬於常見或常成群出現的魚類,現都已變成罕見、稀少甚或是區域性的滅絕;大型的肉食性魚類像海鰻、石鱸、石斑、笛鯛更是明顯,取而代之的是小型的鰕虎、三鰭等隱蔽種,或色彩黯淡不引人注意的小型雀鯛、隆頭魚、粗皮鯛或臭肚魚等等。珊瑚礁魚類魚種的組成和生態系的結構和功能已發生了巨大的改變,原本游魚四出、熱鬧熙攘的海龍宮的場景已不復見。追究原因不外乎是國人難改愛吃、愛養、愛釣的消費習慣,潛水時不經意的踐踏破壞及在礁區潛水打魚。在珊瑚礁區濫捕、廢棄的網具或從事箱網養殖,
許多珊瑚礁魚類都是小個體,連續產卵。另一個共同的特徵是,在生命史的早期,通常有一個浮游擴散階段(Gibson, 1982, 1986;Sale, 1980;Thresher, 1984)。不是卵本身是遠海(pelagic)的-海洋中上層魚類。或者魚類幼體孵化後變成遠海的。這一浮游階段被解釋為一種機制,以避免對珊瑚礁的強烈捕食,實現幼魚的擴散,或使幼魚能夠開發不完整的浮游食物資源。這些共同的特徵不應掩蓋珊瑚礁魚類所顯示的繁殖模式的巨大多樣性(Thresher, 1984)。
在溫帶珊瑚礁上,繁殖通常是季節性的,大多數物種在春季或夏季產卵。一些物種,包括西歐海岸的棉鱈,在冬末春產卵(Fish and Fish, 1989)。
在熱帶珊瑚礁上,有一個範圍,從那些只在一年中有限時間繁殖的物種到那些全年都有一些產卵的物種,儘管在後者,產卵強度通常會達到峰值(Thresher, 1984;Lowe-McConnell, 1987)。牙買加珊瑚礁的魚類在每年的最低氣溫(2月至4月)出現產卵活動的高峰。相比之下,巴拿馬大西洋沿岸的鯧科魚類在12月至3月的旱季產卵量最低(Robertson, 1990年)。
一些礁魚的季節性產卵可以定時,以便在盛行風和洋流的情況下,給中上層海洋魚類的幼魚最好的機會返回珊瑚礁(Johannes, 1981)。然而,巴拿馬礁魚的最大產卵時間和最大補充時間之間幾乎沒有相關性(Robertson, 1990)。
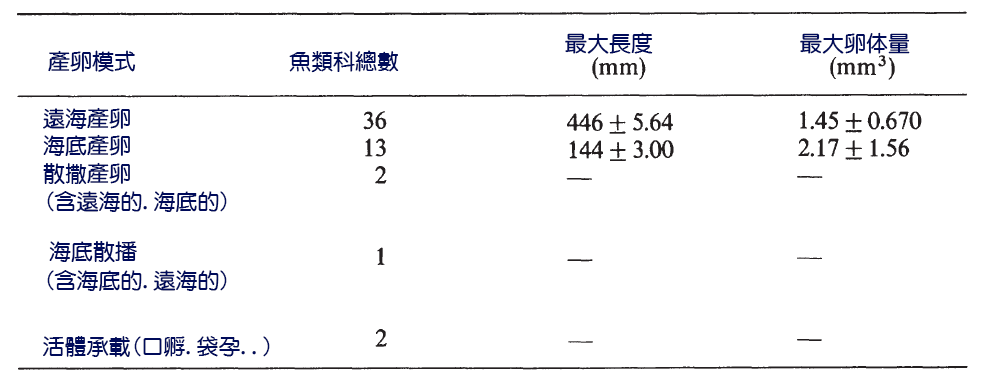
珊瑚礁魚類有五種主要的產卵模式(表7.3),但底棲和遠海產卵是最常見的(Thresher, 1984)。平均而言,遠海產卵者比底棲產卵者多。因此,它們有更高的繁殖力。然而,如果對相同大小的魚進行比較,則底棲和遠海產卵魚種的產卵量和卵大小沒有顯著差異。底棲產卵魚種孵化所需時間比遠海產卵魚種長,而底棲產卵魚種的幼體往往在較高級的發育階段孵化。
許多珊瑚礁魚類保衛領地,在雄性的領地產卵。如果卵是底棲的,它們通常是在不易被湍急的水流沖走的地方產卵,而且卵很少受到捕食。岩縫、石頭的下面和洞是最受歡迎的地方。一些珊瑚礁魚類是築巢者,例如一些溫帶水域的瀨魚。親魚的照顧也很普遍,照顧孩子的父母大多是雄魚,在卵子發育過程中守護並維護環境。天竺鯛科是用嘴孵卵,但它是雄性孵化。在管魚和海馬中,雌魚將卵轉移到雄性魚腹部的育兒袋或凹槽中,在那裡受精,然後發育到甚至超過孵化期。
在其他珊瑚礁魚類聚集在珊瑚礁上,成群的雄性和雌性聚集在一起產卵,卵被釋放到水體中。一些大型魚類,如石斑魚向海遷移,在珊瑚礁向海一側形成大型產卵群。產卵通常發生在落潮和新月和滿月時,潮差最大。這些條件會將卵從礁石中沖走,從而減少棲息在礁石上的魚類和無脊椎動物對它們的捕食,同時也有助於分散魚卵和幼體(Johannes, 1981; Thresher, 1984)。
根據種群密度和種群中魚類的大小分佈,一個物種可能會表現出區域性和群體性產卵(Warner, 1984)。如果成熟魚類的種群密度很高,入侵領地的速度可能會非常快,以致於它們變得站不住腳,並且會成群地產卵。不能保衛領地的小型雄性也會加入產卵群。
雌雄同體,無論是同時性還是連續性,在礁魚中經常被記錄下來。海鱸,Serranidae,是同時存在的雌雄同體,功能性卵巢和睾丸組織存在於同一性腺中(Fischer, 1986)。然而,沒有自我受精。在某些物種中,在產卵過程中,配對者會交替扮演雄性和雌性角色。在塞拉尼德時期,有利於同時雌雄同體進化的選擇性因素尚不清楚。
當兩性的繁殖能力隨大小而增長的速度不同時,順序雌雄同體被認為會進化(Warner, 1984, 1988)。如果雄性的增長速度比雌性快,則預測了從雌性到雄性的變化,如果雌性的增長速度比雄性快,則預測到了雌性與雄性之間的變化。
在礁石生活的魚類,顯示出一種雌雄同體的性別變化,如瀨魚,大雄性能夠隔離一個雌性的後宮,與之交配。因此,作為一個雄魚,生殖的成功取決於獲得一個大的身體尺寸,性別變化從小尺寸的雌魚到雄魚更大的尺寸是有利的。在高種群密度下,小雄魚可能通過參與群體產卵獲得一些繁殖成功。銀蓮花魚形成一夫一妻制配對,但是原雄魚。這可能與體型增加對雌性生殖力的影響有關。在一夫一妻制的一對中,雄魚不會因體型的增加而增加生殖能力,而雌魚則會增加繁殖力。在許多性別變化的物種中,性別的變化是由社會環境的變化引起的,例如,優勢魚、原雌雄同體物種中的雄性和雌性的消失,或者一組魚類的性別比的變化(Shapiro, 1984)。
表7.3 珊瑚礁硬骨魚類產卵模式的分佈,表示為科數,包括顯示特殊照顧管道的物種。魚科可以用多種模式表示。
大多數大陸棚硬骨魚產卵具有漂浮力,大海漂移的魚卵,並沒有提供親魚的照顧。就在產卵之前,卵吸收水分,受精後,卵吸收更多的水,使它們膨脹,變得更具浮力。即使是像鱈魚和比目魚這樣的底棲魚類也會產生遠海魚卵。比目魚的幼體是遠海性的,在其蛻變為幼體前呈側向壓縮的成魚,並在岸邊育雛區生活。
顯然,遠海產卵的一個明顯的例外是大陸棚的軟骨魚類動物,其中許多以胎生的形式表現出親魚的照顧。第二個例外是那些產卵的硬骨魚。包括鯡魚。在北海、大西洋鯡魚產卵在礫石斑塊。太平洋鯡魚,在水下植物上產卵。北大西洋的毛鱗魚(胡瓜魚科),產卵於大陸棚底層(深約200米),有些種群在海灘潮間帶的礫石中產卵。
大陸棚魚類是典型的海洋魚類。有些像鳳尾魚是連續產卵的,而其他一些像鱈魚、鯡魚和比目魚都是同時期一次產卵地。連續產卵與相對較長的繁殖季節有關。然而,即使是那些同時一次產卵的物種也可以延長繁殖季節,在一年中不同的時間產卵。在新斯科舍省聖喬治灣,整個夏季和秋季連續生產鯡魚幼體,而所有鯖魚幼體在6月和7月的2個月內生產,大多數在3周內生產(Lambert and Ware, 1984)。在某些物種中,繁殖的時間非常精確。在鱈魚、鰈魚和鯡魚的北部種群中,產卵日期每年的變化很小(Cushing 1975年)。不同種類的鯡魚在不同的時間產卵。秋季或冬季產卵的魚群產卵量大,但產卵量低,而春季和夏末產卵的魚群產卵量多,但較小。這種卵大小和繁殖力之間的權衡關係到新孵化的幼體所處的攝食條件。當浮游生物較稀疏時,較大的幼體將佔優勢。
對於低緯度的大陸棚魚類,產卵季節往往比高緯度魚類長(Longhurst and Pauly, 1987)。一些物種,例如秘魯鳳尾魚,在產卵活動中表現出明顯的高峰,而其他物種則在一年中呈現出均勻的產卵模式。
產卵的季節性可能與中上層魚類幼體對足夠數量的浮游生物的需求有
關,這些浮游生物的大小在一定的範圍內,可以被攝取。在高緯度地區,有一個明確的浮游生物年生產週期,而在低緯度地區,生產週期沒有那麼強烈的季節性。那
些連續產卵的物種或在繁殖季節產生一系列幼體的物種,在浮游生物產量低的時候,可以降低產生全部幼體的風險,這一生殖策略被稱為”bet-hedging”最佳產卵時機賭注。
許多大陸棚魚類壽命長,因此一個物種的種群包含許多年齡等級。由於幼魚的生產期分佈在多個繁殖季節,因此壽命較長的魚類可降低繁殖失敗的風險。 不良的繁殖季節應該由好的季節來平衡。
海洋上層帶的硬骨魚類的繁殖模式與大陸棚魚類相似。它們產生大量的小型、浮力大的遠海魚卵,沒有親魚的照顧。他們是典型的多次繁殖(iteroparous)物種。軟骨魚類通常是胎生的,在很長的時間間隔內產下一些更多的後代。
《三群魚》(1979年,馬歇爾《落入大海》)。首先,深海軟骨魚類有內受精作用。有些,如銀鮫,產卵大,被保護在一個膠囊裡。其他的,像方形鯊魚,是胎生的。一些深海硬骨獸,包括一些蛋嘴獸(蟲科),也是胎生的。其他的底棲魚類,包括溫帶和極地水域的海螺(海螺科),在發育過程中會產生很大的卵,這些卵可能會留在底部或靠近底部。其中一些底棲硬骨魚可能表現出父母的照顧形式,包括嘴巴的孵卵。許多底棲動物、深海硬骨魚和中層硬骨魚都會產卵,這些卵在水體中向上漂浮。因此,幼體孵化到溫躍層以下、溫躍層內或溫躍層以上的更富生產力的水域。在這些較淺水域生長一段時間後,幼魚會沿著水體向下移動到深水區的父母棲息地。
Marshall(1979, 1984)認為,與水體較高的魚類相比,貧窮的中、深海區的魚類骨骼和肌肉系統都有所減少,因此在身體的體細胞成分上節省投資,以最大限度地增加配子的投資。這使得這些魚能夠產生足夠的卵,以彌補當卵向上漂浮和幼體在多產的淺水中發育時不可避免的損失。
在熱帶和亞熱帶水域中占主導地位的小魚,包括燈籠魚,可能只活一到兩年。在高緯度地區,這些小型的中層遷徙者可能存活數年(Childres等人, 1980年)。中、深海區的大型魚類和大型底棲魚類,如鼠尾(巨尾魚科),壽命長,可能是魚類。然而,Childress等人。(1980)認為一些深海魚類在生命後期才達到性成熟,並且可能只繁殖一次。
低中上層和深海區食物供應稀少,導致種群密度低。這就產生了找伴侶的問題。許多中層魚類具有生物發光性,眼睛發育良好,因此視覺線索可能起作用。深海魚類的眼睛通常會變小,也可以使用其他的感覺方式,如嗅覺。角質釣魚魚表現出一種深刻的性二型性。雄性達到性成熟時的體型比雌性(矮小雄性)小得多,眼睛和嗅覺器官發育良好。雌魚在蛻變為成蟲時,會呈現出球狀的形狀,誘餌發育良好。侏儒雄性有大的睾丸,當它們成熟時,它們會發展出鉗子狀的下顎,在交配時,它們可以依附在雌性上。在一些角質動物中,侏儒雄性會永久性地依附於寄生在她身上的雌性動物,但同時也確保了她始終能夠接觸到配偶。
雌雄同體現象存在於一些深海魚類中,可能是為了適應在低種群密度下尋找配偶的問題。在中層帶,大型肉食性粉蝶龍和無名硬骨獸是同步雌雄同體。綠藻,包括三腳魚,是亞熱帶和熱帶深水區最常見的底棲魚類,也是同步雌雄同體。在這些群體中,自我受精的發生率是未知的。
當考慮到魚類種群豐度及其種群動態的變化時,不同棲息地魚類之間的差異較小,而高繁殖力硬骨魚和低繁殖力軟骨魚類的動態差異更大。種群動態的基本原理適用於這兩個群體。人們使用各種技術來獲得種群豐度的估計值,這是研究種群動態的基礎。這些在1982年被描述。
淨生殖率(Ro)是人口生活史模式成功與否的有用指標。 它可以定義為人口每一代的繁殖率,或者等效地,定義為種群中每位雌性成魚平均生產的雌性幼魚數量(Begon等, 1989)。 (假設受精率不受雄性短缺的限制。)如果平均每個雌性成魚由一個雌性幼魚代替,種群數在數值上將是穩定的,既不會增加也不會減少。 淨生殖率為1。 如果淨生殖率平均小於1,那麼種群數正在減少。 如果該值大於1,則種群數在增加。淨生殖率的數值是由種群中普遍存在的特定年齡的存活率和繁殖力決定的。計算如下:
淨繁殖率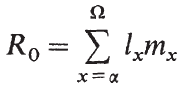
式中,lx是雌性從受精到達到x年齡的概率,mx是以x歲雌性產卵量表示的繁殖力,α=成熟年齡,Ω=種群中可能的最大壽命。
在魚類中,存活率和繁殖力都取決於大小。因此,種群中個體的增長率將通過與個體大小相關的結果來影響Ro。
個體的豐度、增長率和存活率也將決定種群生物量隨時間的變化。生產量是一個群體在規定的時間間隔內合成的組織總量的量度(Mann和Penczak, 1986)。它提供了魚肉對該種群的剝削者—掠食者或食腐動物—的利用率的估計。定義為:
式中,g是比增長率,B是估計產量的時間間隔內種群的平均生物量。種群在任何給定時間的生物量(B)計算如下:
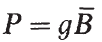
式中,g是比增長率,B是估計產量的時間間隔內種群的平均生物量。種群在任何給定時間的生物量(B)計算如下:
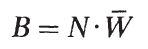
N是種群的豐度,W是當時個體在種群中的平均重量。估計產卵期的產量也必須估計出卵子和精子的貢獻。
一個種群豐度和產量的變化模式將部分反映出種群自身密度對其死亡率、繁殖力和生長的影響的重要性(Begon等人, 1989年)。如果密度的影響使得在高種群密度下,平均Ro小於1(種群減少),而在低種群密度下,平均速率大於1(種群增長),則在沒有其他干擾因素的情況下,種群將在平均Ro為1的密度附近波動(圖7.3)。一個種群在數量減少後,其數量可能會增加,這對漁業很重要。通過從一個種群中捕撈魚類,漁場在死亡率、出生率和增長率方面設定了一個依賴於列車密度的變化,從而補償了漁業的損失。
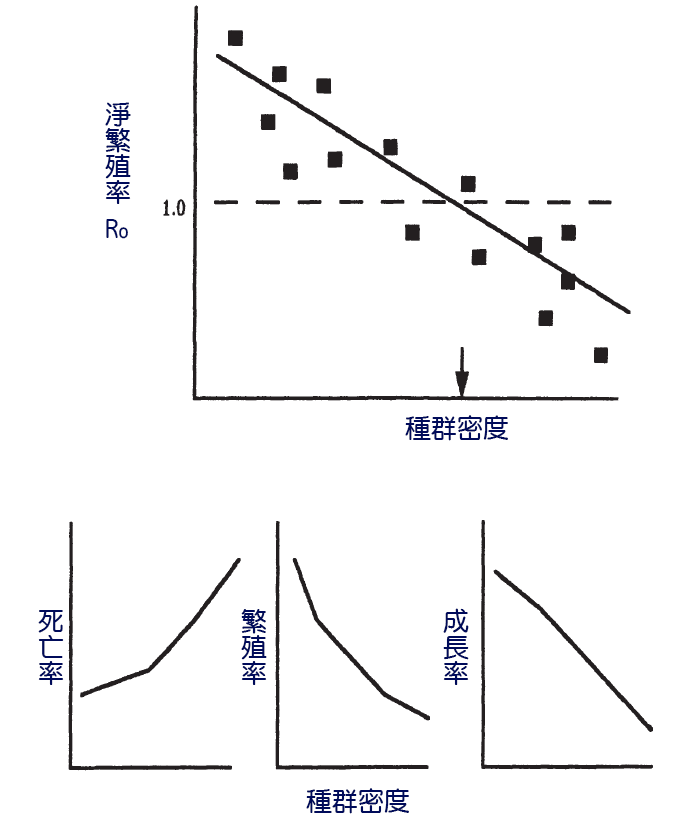
圖7.3 由淨生殖率(Ro)、死亡率、繁殖力和生長的密度依賴關係表示的密度依賴關係。垂直箭頭表示平均Ro等於1.0的平衡種群密度。
儘管Ro的密度依賴效應(通常)趨向於穩定種群的數量豐度,但隨著時間的推移,魚類種群會陷入絕望。漁業生物學家花了大量的精力來描述魚群的招募關係,以一種對管理漁業有用的方式。Ricker(1954)和Beverton and Holt(1957)的研究尤其具有影響力。
許多種群的第二個特點是存在佔優勢的年級或佇列。當繁殖季節繁殖出異常多的幼魚時,通常是由於不完全瞭解的原因而產生的。在長壽魚中,一個優勢年級的存在通常在它第一次出現後的許多年內都能被檢測到,因為在任何年齡段,它在總種群中所占的比例都高於通常預期的那個年齡段的魚。一個著名的例子是1904年的大西洋鯡魚, 1922年在人群中仍然可以檢測到(Nikolskii 1969)。1963年北海普拉斯班的招募人數比每年平均招募人數多出200%(Rothschild 1986年)。在已開發的魚類種群中,出現優勢年級通常可以獲得更高的產量。
不確定的種群-再利用關係和優勢年類現象突出了理解影響產卵和性成熟開始之間生存的過程的重要性。生命最初幾個月發生的事情通常是至關重要的。捕食和食物充足,以及溫度和水運動的影響,可能是影響幼魚存活的主要因素(Bailey和Houde, 1989)。這些因素可能以複雜的方式相互作用。在高食物水準下,魚可以生長得更快,因此不易被捕食。如果有足夠的食物,生長速度往往會隨著溫度的升高而增加,但是更高的溫度也會導致更高的新陳代謝率和更饑餓的捕食者。
在某些物種中,卵黃被吸收後開始獨立攝食可能是一個關鍵時期。如果幼魚適宜食物的濃度不足,魚就會變弱,變得更容易被捕食或挨餓。Cushing(1990)在他的匹配錯配假說中擴展了這一觀點。這預示著,當仔魚開始攝食的時間與生產適合幼魚和幼魚的食物時,攝食、生長和存活都會很好。這將使那群魚得到很好的招募。如果幼體在食物稀少的時候開始進食,那麼從該群體中招募的數量將很低。
儘管有一些間接證據(Rothschild, 1986;Cushing, 1990),以一種能夠對其死亡率和增長率提供良好定量估計的方法對海洋中的幼魚和幼魚進行取樣的困難使得很難檢驗臨界期和“匹配錯配”的假設(Rothschild, 1986;Cushing, 1990)。加拿大東海岸附近的海岬海在年級上表現出較大的年際波動力量。r這裡年級強度與兩個非生物環境變數之間有很好的相關性:孵化後立即出現的陸上風頻率,以及隨後幼體漂流期間的水溫(Leggett等人, 1984年)。這是為數不多的例子之一,其中一個令人信服的因果關係可以給出解釋。陸上的風把溫暖的地表水吹到海岬魚 產卵的海灘上。溫度的升高刺激了幼體角鬥魚從礫石中冒出,這一過程受到湍急水流的干擾。這種溫暖的地表水攜帶的潛在幼體捕食者的密度比離岸風吹起時上升的 更深、更冷的水要低。溫暖的海水中還有一個浮游生物群落,其中含有更適合幼體的獵物。在有足夠食物的溫暖水域中,幼體可以更快地生長,這將減少它們受到某 些體型選擇性捕食者的威脅的時間。
關鍵時期最明顯的例子來自於對英格蘭湖區溪流中鱒魚Salmo trutta種群生物學的研究。Elliott(1989)的這項研究是硬骨魚種群動態研究的一個突出例子。主要分析是鮭魚的溯河產卵種群(圖8.4)。孵化後,魚在溪流中待2年,然後才遷移到河口或海洋。返回產卵前在海裡待的時間從不到一年到兩年多不等。這種魚群是硬骨魚的典型特徵,因為產卵魚群的大小只是魚群數量的一個很差的預測因數(圖7.5)。然而,分析表明,無論產卵的數量如何,在生命的第一個夏天結束時,幼仔的數量是相似的。埋在岩縫中的卵存活率很高,但從岩縫中冒出後,頭幾個月的死亡率強烈依賴於密度。仔細取樣表明,在幼體成長後有一個關鍵時期,在此期間死亡率依賴於密度。在這個關鍵時期之後,死亡率不再依賴於密度。臨界期的長度在30到70天 之間變化,並且本身是密度依賴的。週期越短,卵密度越高。在該群體中,種群的主要密度依賴性調節發生在出苗後的關鍵時期。出苗後的生長速度與密度無關,但 在臨界期,無論是大鱒魚還是小鱒魚,在高密度下的存活率都低於中型鱒魚。鱒魚生存的關鍵是在關鍵時期佔據覓食區域的能力。在這一時期的開始,往往有大量沒 有領地的鱒魚,但到了這一時期結束時,數量很少。
鮭魚生活史的特點,包括卵的埋藏、幼魚從沙礫中冒出時的體型較大以及隨後的取食區域,使這項研究的結果很難概括為散佈其卵的硬骨魚,其幼體孵化的尺寸較小且脆弱。對鮭魚種群動態的詳細研究強調了早期種群動態的研究價值。
鳳尾魚似乎已交替成為主要的中上層浮游動物。在歐洲西北部,鯡魚的數量呈現出極大的豐盛時期,它們支持了繁榮的漁業和其他短缺時期。在較短的時間尺度上,庫欣(1990)提供了證據,證明了西北大西洋幾個底棲種群的不良招募與大量異常涼爽和新鮮的水橫穿大西洋之間可能存在聯繫。
只有更好地理解食物水準、非生物因素和生物相互作用對生長、死亡和生殖的影響,才能理解氣候條件對種群動態的影響。
由於軟骨魚類在經濟上的重要性要低得多,它們的種群動態比硬骨魚 受到的關注要少。它們長壽、生長緩慢、成熟期晚、妊娠期長、繁殖力低的生活史特徵表明,它們對種群密度變化的反應會比大多數硬骨魚慢得多。因此,它們對漁 業損失的補償能力可能不如硬骨魚,只能承受較輕的捕撈壓力。在愛爾蘭海,無鰭鰩科raiabatis,由於捕魚壓力而瀕臨滅絕(Brander, 1988)。在泰國灣,底棲漁業捕撈的鰩魚從1963年的每拖網捕魚小時15.8公斤下降到1982年的0.1公斤(Pauly, 1988)。
產量是指一個種群在一定時間內合成的魚組織的總量,包括在這段時間內死亡的個體合成的魚組織。因此,它是不同生物學過程的結果:生長、死亡和繁殖。對於個別淡水物種,估計產量在0.01-156.4 g/㎡/年之間。產量與生物量之比P/B的估計值在0.2-8.5之間(Mann和Penczak, 1986)。
即使在一個種群中,年產量也可能相差很大。雖然生產價值大體上反映了一個種群對其他消費者的潛在收益率,但這些價值本身並不能深入瞭解種群在社區運作中的作用。在水生生物群落中,許多相互作用取決於相互作用體的大小。重要的問題與其說是生產總量,不如說是生產在種群中不同規模階層之間的分配方式。在增長率和死亡率都很高的情況下,最年輕年齡階層的產量往往占總產量的很高比例。然而,這方面的研究仍處於初級階段。
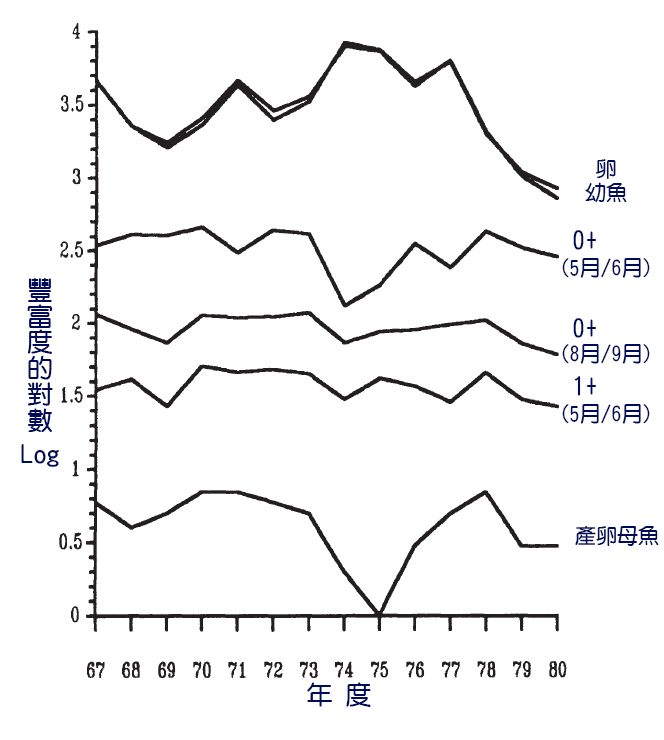
圖7.4: 溯河鱒魚種群動態,顯示了各年的豐度:(i)卵;(ii)幼魚;(iii)0+(5/6月)parr;(iv)O+(8月/9月)parr;(v)1+(5月/6月)parr和(vi)成年產卵者。(年份等級是指從礫石中出現的年份,而不是卵的產卵年份。)每個階段的豐度(每60平方米)以IOg10組織給出。Elliott (1985).

圖7.5 上圖所示溯河鱒魚種群的種群招募關係。存量表示為一年開始時出現的魚卵數量,新魚數量表示為當年生產的魚卵數量(每60平方米1個卵)。虛線是等額替換線,也就是說,新魚卵=種群招募。注意種群招募和新魚之間的弱相關。此外,有證據表明,在低密度下,新魚卵超過種群招募,在高密度時,種群招募超過新魚卵。Elliott
(1985).